

400-998-5282
专注多肽 服务科研
400-998-5282
专注多肽 服务科研
这是一种易溶、特异且敏感的胰凝乳蛋白酶和人胰弹性蛋白酶底物。它也能被组织蛋白酶G和糜蛋白酶水解。
编号:111237
CAS号:70967-97-4
单字母:Suc-AAPF-pNA
| 编号: | 111237 |
| 中文名称: | 组织蛋白酶G基质I:Suc-Ala-Ala-Pro-Phe-pNA |
| CAS号: | 70967-97-4 |
| 单字母: | Suc-AAPF-pNA |
| 三字母: | Suc N端琥珀酰化,琥珀酰基是一种极性的N端修饰,对应于脱氨基天冬氨酸(deamino-Asp),常被掺入羧肽酶底物中。 -Ala丙氨酸:alanine。L-丙氨酸的系统命名为(2S)-氨基丙酸,是编码氨基酸,也叫L-α-丙氨酸。符号:A,Ala。D-丙氨酸存在于多种细菌细胞壁的糖肽中。β-丙氨酸是维生素泛酸和辅酶A的组分。 -Ala丙氨酸:alanine。L-丙氨酸的系统命名为(2S)-氨基丙酸,是编码氨基酸,也叫L-α-丙氨酸。符号:A,Ala。D-丙氨酸存在于多种细菌细胞壁的糖肽中。β-丙氨酸是维生素泛酸和辅酶A的组分。 -ProL-脯氨酸:proline。系统命名为吡咯烷-(2S)-羧酸。为亚氨基酸。是编码氨基酸。在肽链中有特殊作用,如易形成顺式的肽键等。符号:P,Pro。 -PheL-苯丙氨酸:phenylalanine。系统命名为(2S)-氨基-3-苯基丙酸。是编码氨基酸。是哺乳动物的必需氨基酸。符号:F,Phe。 -pNA对硝基苯胺 |
| 氨基酸个数: | 4 |
| 分子式: | C30H36O9N6 |
| 平均分子量: | 624.64 |
| 精确分子量: | 624.25 |
| 等电点(PI): | - |
| pH=7.0时的净电荷数: | - |
| 平均亲水性: | -1.1666666666667 |
| 疏水性值: | 1.2 |
| 外观与性状: | 白色粉末状固体 |
| 消光系数: | - |
| 来源: | 人工化学合成,仅限科学研究使用,不得用于人体。 |
| 纯度: | 95%、98% |
| 盐体系: | 可选TFA、HAc、HCl或其它 |
| 储存条件: | 负80℃至负20℃ |
| 标签: | 细胞凋亡肽(Apoptosis Peptides) pNA修饰肽 Suc修饰肽 |
| 参考文献(References): | Nakajima, K., and Powers, J.C. The Journal of Biological Chemistry 254(10), 4027-4032 (1979). Ermolieff, J., Boudier, C., Laine, A., et al. The Journal of Biological Chemistry 269(47), 29502-29508 (1994). Nakakubo, H., Fukuyama, H., Nakajima, M., et al. Yeast 16(4), 315-323 (2000). |
这是一种易溶、特异且敏感的胰凝乳蛋白酶和人胰弹性蛋白酶底物。它也能被组织蛋白酶G和糜蛋白酶水解。此外,它还是FK-506结合蛋白(FKBPs,也称为巨噬细胞素)和亲环素的标准底物,这些蛋白属于肽基脯氨酰顺反异构酶(PPIases)类。因此,Suc-AAPF-pNA已被用于PPIase活性的非偶联无蛋白酶检测。
A readily soluble, specific and sensitive substrate for chymotrypsin and human pancreatic elastase. It is also hydrolyzed by cathepsin G and chymase. Furthermore it is the standard substrate for FK-506 binding proteins (FKBPs, also called macrophilins) and cyclophilins, which belong to the group of peptidyl prolyl cis-trans isomerases (PPIases). Thus, Suc-AAPF-pNA has been used for an uncoupled protease-free assay of PPIase activity.
N-琥珀酰-L-丙氨酰-L-丙氨酰-L-脯氨酰-L-苯丙氨酸4-硝基苯胺是肽基脯氨酰异构酶、胰凝乳蛋白酶和人白细胞组织蛋白酶G的比色底物,但不与弹性蛋白酶反应。
N-Succinyl-L-alanyl-L-alanyl-L-prolyl-L-phenylalanine 4-nitroanilide is a colorimetric substrate for peptidyl-prolyl isomerase, chymotrypsin and human leukocyte cathepsin G but not reactive with elastase.
Suc-AAPF-pNA是一种对人白细胞组织蛋白酶G敏感的显色底物,不与弹性蛋白酶反应。它可被组织蛋白酶G(Km = 1.7 mM)、枯草杆菌蛋白酶、胰凝乳蛋白酶(Km = 60 µM)、糜蛋白酶(Km = 4 mM)和亲环素裂解。在405-410 nm处监测对硝基苯胺的释放。该底物可用于抑制剂筛选和动力学分析。
Suc-AAPF-pNA is a sensitive chromogenic substrate for human leukocyte cathepsin G that is not reactive with elastasecan. It can be cleaved by cathepsin G (Km = 1.7 mM), subtilisins, chymotrypsin (Km = 60 µM), chymase (Km = 4 mM), and cyclophilin. Release of p-nitroanilide is monitored at 405-410 nm. This substrate can be used for inhibitor screening and kinetic analysis.
琥珀酰基 - 丙氨酸 - 丙氨酸 - 脯氨酸 - 苯丙氨酸 - 对硝基苯胺在切割位点带有苯丙氨酸,靶向糜蛋白酶样蛋白酶。序列背景在脯氨酸周围稳定转角结构。生色输出可进行定量酶分析。研究人员将其用于动力学检测与底物图谱分析。
Suc-Ala-Ala-Pro-Phe-pNA features a phenylalanine at the cleavage site to target chymotrypsin-like proteases. Sequence context stabilizes a turn around proline. Chromogenic output allows quantitative enzyme analysis. Researchers use it in kinetic assays and substrate mapping.
Definition
Apoptosis or programmed cell death is a normal component of the development and health of multicellular organisms. Cells die in response to a variety of stimuli and during apoptosis they do so in a controlled, regulated fashion.
Discovery
In 1885, Flemming W described the process of programmed cell death. John Kerr's discovery, in late 1960s, initially called "shrinkage necrosis" but which he later renamed "apoptosis", came about when his attention was caught by a curious form of liver cell death during his studies of acute liver injury in rats 1,2. Kerr in 1972 proposed the term apoptosis is for mechanism of controlled cell deletion, which appears to play a complementary but opposite role to mitosis in the regulation of animal cell populations. Its morphological features suggest that it is an active, inherently programmed phenomenon, and it has been shown that it can be initiated or inhibited by a variety of environmental stimuli, both physiological and pathological 3.
Structural Characteristics
Heterodimerization between members of the Bcl-2 family of proteins is a key event in the regulation of programmed cell death. The molecular basis for heterodimer formation was investigated by determination of the solution structure of a complex between the survival protein Bcl-xL and the death-promoting region of the Bcl-2-related protein Bak. The structure and binding affinities of mutant Bak peptides indicate that the Bak peptide adopts an amphipathic helix that interacts with Bcl-xL through hydrophobic and electrostatic interactions. Mutations in full-length Bak that disrupt either type of interaction inhibit the ability of Bak to heterodimerize with Bcl-xL 4.
The structure of the 16–amino acid peptide complexed with a biologically active deletion mutant of Bcl-xL was determined by nuclear magnetic resonance spectroscopy (NMR). The structure was determined from a total of 2813 NMR-derived restraints and is well defined by the NMR data. The Bak peptide forms a helix when complexed to Bcl-xL. The COOH terminal portion of the Bak peptide interacts predominantly with residues in the BH2 and BH3 regions. Melanoma inhibitor of apoptosis (ML-IAP) is a potent anti-apoptotic protein that is upregulated in a number of melanoma cell lines but not expressed in most normal adult tissues. Overexpression of IAP proteins, such as ML-IAP or the ubiquitously expressed X-chromosome-linked IAP (XIAP), in human cancers has been shown to suppress apoptosis induced by a variety of stimuli. X-ray crystal structures of ML-IAP-BIR in complex with Smac- and phage-derived peptides, together with peptide structure−activity-relationship data, indicate that the peptides can be modified to provide increased binding affinity and selectivity for ML-IAP-BIR relative to XIAP-BIR3 5.
Mode of Action
Upon receiving specific signals instructing the cells to undergo apoptosis a number of distinctive changes occur in the cell. Families of proteins known as caspases are typically activated in the early stages of apoptosis. These proteins breakdown or cleave key cellular components that are required for normal cellular function including structural proteins in the cytoskeleton and nuclear proteins such as DNA repair enzymes. The caspases can also activate other degradative enzymes such as DNases, which begin to cleave the DNA in the nucleus.
Apoptotic cells display distinctive morphology during the apoptotic process. Typically, the cell begins to shrink following the cleavage of lamins and actin filaments in the cytoskeleton. The breakdown of chromatin in the nucleus often leads to nuclear condensation and in many cases the nuclei of apoptotic cells take on a "horse-shoe" like appearance. Cells continue to shrink, packaging themselves into a form that allows for their removal by macrophages. There are a number of mechanisms through which apoptosis can be induced in cells. The sensitivity of cells to any of these stimuli can vary depending on a number of factors such as the expression of pro- and anti-apoptotic proteins (eg. the Bcl-2 proteins or the Inhibitor of Apoptosis Proteins), the severity of the stimulus and the stage of the cell cycle. The Bcl-2 family of proteins plays a central role in the regulation of apoptotic cell death induced by a wide variety of stimuli. Some proteins within this family, including Bcl-2 and Bcl-xL, inhibit programmed cell death, and others, such as Bax and Bak, can promote apoptosis 6, 7.
Functions
For development, Apoptosis is as needed for proper development as mitosis is. Examples: The resorption of the tadpole tail at the time of its metamorphosis into a frog occurs by apoptosis.
Integrity of the organism, Apoptosis is needed to destroy cells that represent a threat to the integrity of the organism. Examples: Cells infected with viruses8.
Cells of the immune system, as cell-mediated immune responses wane, the effector cells must be removed to prevent them from attacking body constituents. CTLs induce apoptosis in each other and even in themselves 9.
Cells with DNA damage, damage to its genome can cause a cell to disrupt proper embryonic development leading to birth defects to become cancerous.
References
1. Kerr JF (1965). A histochemical study of hypertrophy and ischaemic injury of rat liver with special reference to changes in lysosomes. Journal of Pathology and Bacteriology, 90(90):419-435.
2. Kerr JF, Wyllie AH, Currie AR (1972). Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer., 26(4):239-257.
3. O'Rourke MG, Ellem KA (2000). John Kerr and apoptosis. Med. J. Aust., 173(11-12): 616-617.
4. Franklin MC, Kadkhodayan S, Ackerly H, Alexandru D, Distefano MD, Elliott LO, Flygare JA, Mausisa G, Okawa DC, Ong D, Vucic D, Deshayes K, Fairbrother WJ (2003). Structure and function analysis of peptide antagonists of melanoma inhibitor of apoptosis (ML-IAP). Biochemistry, 42(27):8223-8231.
5. Sattler M, Liang H, Nettesheim D, Meadows RP, Harlan JE, Eberstadt M, Yoon HS, Shuker SB, Chang BS, Minn AJ, Thompson CB, Fesik SW (1997). Structure of bcl-xl-bak peptide complex: recognition between regulators of apoptosis. Science, 275(5302):983-986.
6. Hanada M, Aimé-Sempé C, Sato T, Reed JC (1995). Structure-function analysis of Bcl-2 protein. Identification of conserved domains important for homodimerization with Bcl-2 and heterodimerization with Bax. J. Biol. Chem., 270(20):11962-11969.
7. Cheng EHY, Levine B, Boise LH, Thompson CB, Hardwic JM (1996). Bax-independent inhibition of apoptosis by Bcl-xL.Nature, 379:554-556.
8. Alimonti JB, Ball TB, Fowke KR (2003). Mechanisms of CD4+ T lymphocyte cell death in human immunodeficiency virus infection and AIDS. J Gen Virology., 84(84): 1649-1661.
9. Werlen G, Hausmann B, Naeher D, Palmer E (2003). Signaling life and death in the thymus: timing is everything. Science. 299(5614):1859-1863.
| DOI | 名称 | |
|---|---|---|
| 10.1074/jbc.M411482200 | Granzyme M is a regulatory protease that inactivates proteinase inhibitor 9, an endogenous inhibitor of granzyme B | 下载 |
| 10.1016/j.febslet.2006.04.087 | Comparative analysis of enzyme activities and mRNA levels of peptidyl prolyl cis/trans isomerases in various organs of wild type and Pin1-/- mice | 下载 |
| 10.1016/s0014-5793(01)03053-8 | Functional analysis of the propeptides of subtilisin E and aqualysin I as intramolecular chaperones | 下载 |
| 10.1021/bi015650+ | Heterogeneity in serpin-protease complexes as demonstrated by differences in the mechanism of complex breakdown | 下载 |
| 10.1016/s0014-5793(02)02242-1 | Evidence for diversity of substrate specificity among members of the chymase family of serine proteases | 下载 |
| 10.1110/ps.3510102 | Analysis of serine proteinase-inhibitor interaction by alanine shaving | 下载 |
| 10.1016/S0003-9861(02)00054-1 | Inhibition of cathepsin G by 2-amino-3,1-benzoxazin-4-ones: kinetic investigations and docking studies | 下载 |
| 10.1021/jm020865i | Synthesis and evaluation of Glypsi(PO(2)R-N)Pro-containing pseudopeptides as novel inhibitors of the human cyclophilin hCyp-18 | 下载 |
| 10.1074/jbc.M205517200 | Functional analysis of Toxoplasma gondii protease inhibitor 1 | 下载 |
| 10.1074/jbc.M301512200 | Extended substrate specificity of rat mast cell protease 5, a rodent alpha-chymase with elastase-like primary specificity | 下载 |
| 10.1021/bi034773m | Do enzymes change the nature of transition states? Mapping the transition state for general acid-base catalysis of a serine protease | 下载 |
| 10.1046/j.1432-1033.2003.03809.x | Activated transglutaminase from Streptomyces mobaraensis is processed by a tripeptidyl aminopeptidase in the final step | 下载 |
| 10.1016/j.molbiopara.2003.10.003 | Molecular cloning, expression and characterization of a serine proteinase inhibitor gene from Entamoeba histolytica | 下载 |
| 10.1111/j.1365-294X.2004.02381.x | Impact of oilseed rape expressing the insecticidal serine protease inhibitor, mustard trypsin inhibitor-2 on the beneficial predator Pterostichus madidus | 下载 |
| 10.1016/j.febslet.2005.08.056 | Differences between human proteinase 3 and neutrophil elastase and their murine homologues are relevant for murine model experiments | 下载 |
| 10.1074/jbc.M507760200 | The protein phosphatase 2A phosphatase activator is a novel peptidyl-prolyl cis/trans-isomerase | 下载 |
| 10.1002/jps.20512 | Effect of cyclodextrins on alpha-chymotrypsin stability and loading in PLGA microspheres upon S/O/W encapsulation | 下载 |
| 10.1016/j.bbapap.2006.04.015 | Compatible solutes as protectants for zymogens against proteolysis | 下载 |
| 10.1515/znc-2006-5-623 | Catalytic efficiencies of alkaline proteinases from microorganisms | 下载 |
| 10.1021/bi0608820 | Thermodynamics of phosphopeptide binding to the human peptidyl prolyl cis/trans isomerase Pin1 | 下载 |
| 10.1016/j.jmb.2007.02.032 | Structural and mechanistic exploration of acid resistance: kinetic stability facilitates evolution of extremophilic behavior | 下载 |
| 10.1074/jbc.M702615200 | Inhibition of the activation of multiple serine proteases with a cathepsin C inhibitor requires sustained exposure to prevent pro-enzyme processing | 下载 |
| 10.1016/j.bbrc.2007.05.124 | Identification and characterization of a novel and functional murine Pin1 isoform | 下载 |
| 10.1074/jbc.M707157200 | Structural basis for elastolytic substrate specificity in rodent alpha-chymases | 下载 |
| 10.1124/mol.108.045682 | In vivo inhibition of serine protease processing requires a high fractional inhibition of cathepsin C | 下载 |
| 10.1111/j.1742-4658.2008.06333.x | Functional domains of the human epididymal protease inhibitor, eppin | 下载 |
| 10.1111/j.1742-4658.2008.06381.x | The crystal structure of human WD40 repeat-containing peptidylprolyl isomerase (PPWD1) | 下载 |
| 10.1515/BC.2008.153 | Cytotoxic and peptidase inhibitory activities of selected non-hepatotoxic cyclic peptides from cyanobacteria | 下载 |
| 10.1074/jbc.M803041200 | Activation mechanism of recombinant Der p 3 allergen zymogen: contribution of cysteine protease Der p 1 and effect of propeptide glycosylation | 下载 |
| 10.1002/cbic.200800415 | New peptolides from the cyanobacterium Nostoc insulare as selective and potent inhibitors of human leukocyte elastase | 下载 |
| 10.3390/molecules14010378 | 2-Amino- and 2-alkylthio-4H-3,1-benzothiazin-4-ones: synthesis, interconversion and enzyme inhibitory activities | 下载 |
| 10.1186/1472-6807-9-17 | Solution structure of the parvulin-type PPIase domain of Staphylococcus aureus PrsA--implications for the catalytic mechanism of parvulins | 下载 |
| 10.1021/bi9007287 | Isoform-specific inhibition of cyclophilins | 下载 |
| 10.1016/j.bbamcr.2010.05.006 | Identification of an atypical peptidyl-prolyl cis/trans isomerase from trypanosomatids | 下载 |
| 10.1016/j.bcp.2010.06.014 | Potency variation of small-molecule chymase inhibitors across species | 下载 |
| 10.1371/journal.pbio.1000439 | Structural and biochemical characterization of the human cyclophilin family of peptidyl-prolyl isomerases | 下载 |
| 10.1186/1472-6750-10-57 | Moisture-induced solid state instabilities in alpha-chymotrypsin and their reduction through chemical glycosylation | 下载 |
| 10.1111/j.1567-1364.2011.00749.x | Recombinant expression of ShPI-1A, a non-specific BPTI-Kunitz-type inhibitor, and its protection effect on proteolytic degradation of recombinant human miniproinsulin expressed in Pichia pastoris | 下载 |
| 10.1371/journal.pone.0037921 | Identification and characterisation of a novel acylpeptide hydrolase from Sulfolobus solfataricus: structural and functional insights | 下载 |
| 10.1016/j.rinphs.2012.11.001 | Two-step nanoprecipitation for the production of protein-loaded PLGA nanospheres | 下载 |
| 10.1016/j.rinphs.2012.08.001 | Glycosylation improves α-chymotrypsin stability upon encapsulation in poly(lactic-co-glycolic)acid microspheres | 下载 |
| 10.1002/bip.22184 | Hybrid analogues of SFTI-1 modified in P₁ position by β- and γ-amino acids and N-substituted β-alanines | 下载 |
| 10.1016/j.fob.2012.07.001 | The role of substrate specificity and metal binding in defining the activity and structure of an intracellular subtilisin | 下载 |
| 10.1111/febs.12542 | Investigation of peptide splicing using two-peptide-chain analogs of trypsin inhibitor SFTI-1 | 下载 |
| 10.1111/febs.12610 | A novel class of bifunctional acylpeptide hydrolases--potential role in the antioxidant defense systems of the Antarctic fish Trematomus bernacchii | 下载 |
| 10.1016/j.bbagen.2013.11.017 | Der p 1 is the primary activator of Der p 3, Der p 6 and Der p 9 the proteolytic allergens produced by the house dust mite Dermatophagoides pteronyssinus | 下载 |
| 10.3390/toxins6030850 | Elapid snake venom analyses show the specificity of the peptide composition at the level of genera Naja and Notechis | 下载 |
| 10.1016/j.bbapap.2014.08.011 | The role of salt bridges on the temperature adaptation of aqualysin I, a thermostable subtilisin-like proteinase | 下载 |
| 10.3390/toxins6092719 | Biochemical characterization of the SPATE members EspPα and EspI | 下载 |
| 10.1021/acsami.5b09301 | Stable and Simple Immobilization of Proteinase K Inside Glass Tubes and Microfluidic Channels | 下载 |
| 10.1016/s0968-0896(00)00281-9 | 6-Acylamino-2-1[(ethylsulfonyl)oxy]-1H-isoindole-1,3-diones mechanism-based inhibitors of human leukocyte elastase and cathepsin G: effect of chirality in the 6-acylamino substituent on inhibitory potency and selectivity | 下载 |
| 10.1186/1471-2172-3-2 | Enhanced immunogenicity of a functional enzyme by T cell epitope modification | 下载 |
| 10.1046/j.1432-1033.2003.03703.x | Transglutaminase from Streptomyces mobaraensis is activated by an endogenous metalloprotease | 下载 |
| 10.1016/j.molbiopara.2007.07.013 | Biochemical properties of an intracellular serpin from Echinococcus multilocularis | 下载 |
| 10.1021/ja807827k | Cleavage of four carbon-carbon bonds during biosynthesis of the griseorhodin a spiroketal pharmacophore | 下载 |
| 10.1021/bm200709r | Monitoring the in vitro enzyme-mediated degradation of degradable poly(ester amide) for controlled drug delivery by LC-ToF-MS | 下载 |
| 10.1055/s-0032-1321846 | Effects of milk casein derived tripeptides on endothelial enzymes in vitro; a study with synthetic tripeptides | 下载 |
| 10.1074/jbc.M113.476580 | A novel family of soluble minimal scaffolds provides structural insight into the catalytic domains of integral membrane metallopeptidases | 下载 |
| 10.1038/srep02985 | Functional role of the flexible N-terminal extension of FKBP38 in catalysis | 下载 |
| 10.1042/BCJ20160098 | Inhibition of Aβ(1-40) fibril formation by cyclophilins | 下载 |
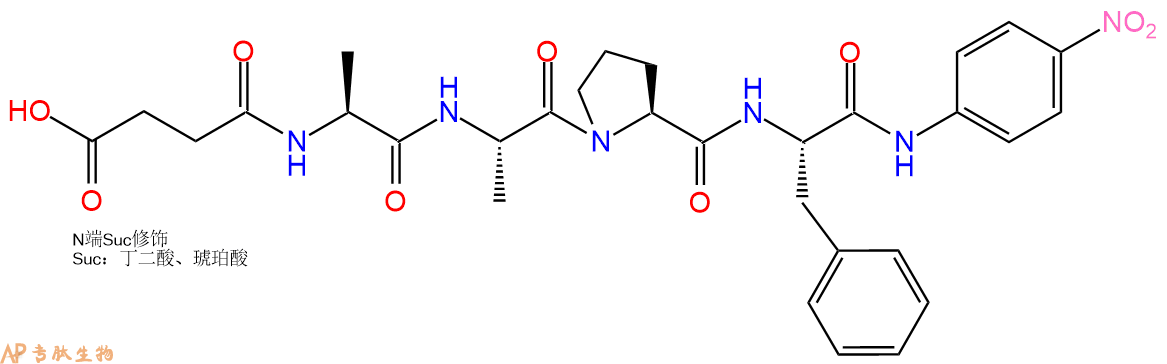
多肽Suc-Ala-Ala-Pro-Phe-pNA的合成步骤:
1、合成CTC树脂:称取2.56g CTC Resin(如初始取代度约为0.57mmol/g)和1.75mmol Fmoc-Phe-OH于反应器中,加入适量DCM溶解氨基酸(需要注意,此时CTC树脂体积会增大好几倍,避免DCM溶液过少),再加入4.38mmol DIPEA(Mw:129.1,d:0.740g/ml),反应2-3小时后,可不抽滤溶液,直接加入1ml的HPLC级甲醇,封端半小时。依次用DMF洗涤2次,甲醇洗涤1次,DCM洗涤一次,甲醇洗涤一次,DCM洗涤一次,DMF洗涤2次(这里使用甲醇和DCM交替洗涤,是为了更好地去除其他溶质,有利于后续反应)。得到 Fmoc-Phe-CTC Resin。结构图如下:
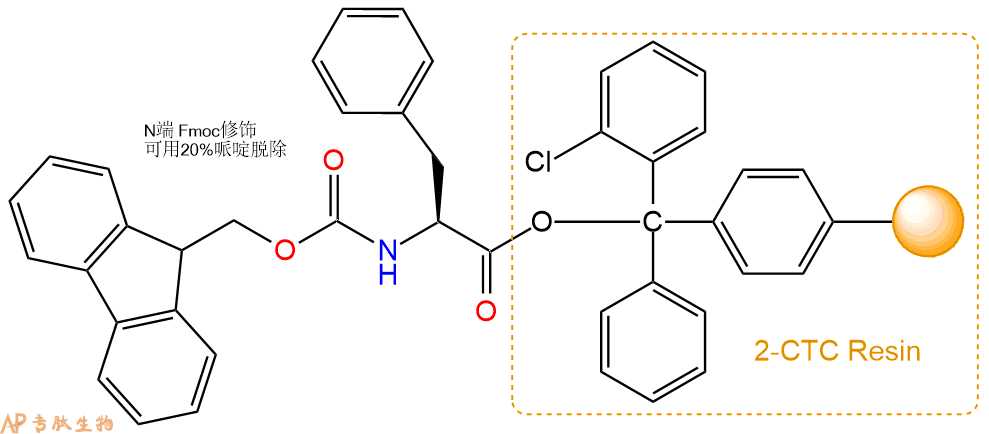
2、脱Fmoc:加3倍树脂体积的20%Pip/DMF溶液,鼓氮气30分钟,然后2倍树脂体积的DMF 洗涤5次。得到 H2N-Phe-CTC Resin 。(此步骤脱除Fmoc基团,茚三酮检测为蓝色,Pip为哌啶)。结构图如下:
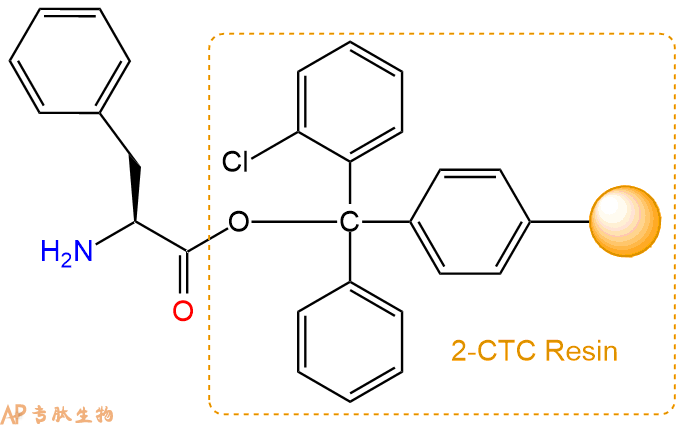
3、缩合:取4.38mmol Fmoc-Pro-OH 氨基酸,加入到上述树脂里,加适当DMF溶解氨基酸,再依次加入8.76mmol DIPEA,4.16mmol HBTU。反应30分钟后,取小样洗涤,茚三酮检测为无色。用2倍树脂体积的DMF 洗涤3次树脂。(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。得到Fmoc-Pro-Phe-CTC Resin。氨基酸:DIPEA:HBTU:树脂=3:6:2.85:1(摩尔比)。结构图如下:
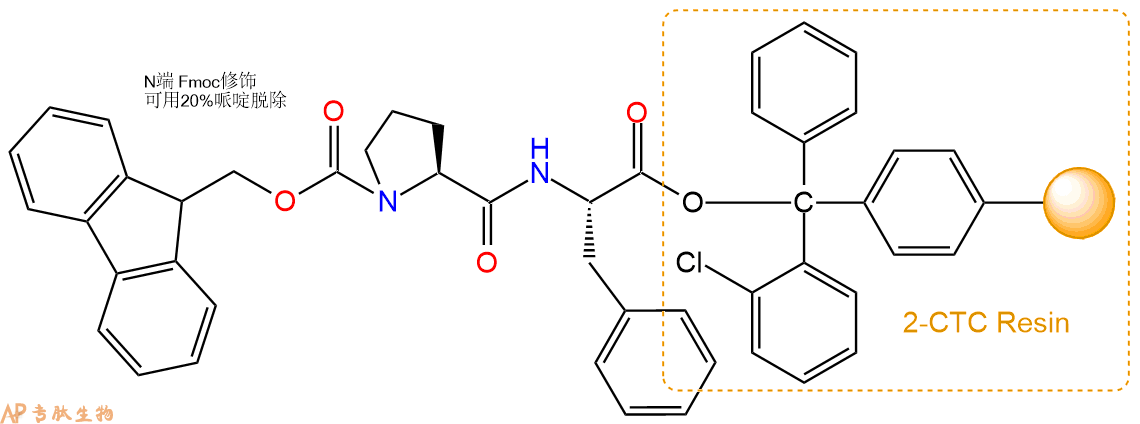
4、依次循环步骤二、步骤三,依次得到
H2N-Pro-Phe-CTC Resin
Fmoc-Ala-Pro-Phe-CTC Resin
H2N-Ala-Pro-Phe-CTC Resin
Fmoc-Ala-Ala-Pro-Phe-CTC Resin
以上中间结构,均可在专肽生物多肽计算器-多肽结构计算器中,一键画出。
最后再经过步骤二得到 H2N-Ala-Ala-Pro-Phe-CTC Resin,结构如下:
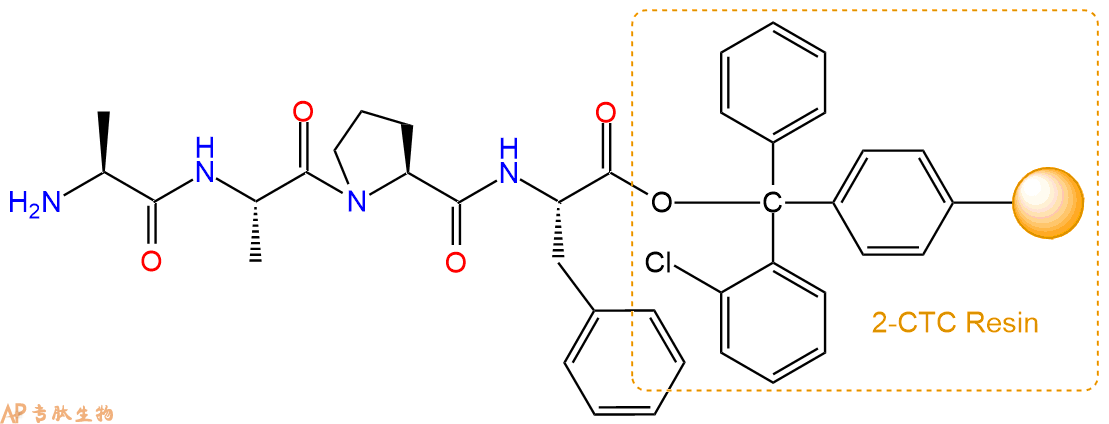
5、丁二酸反应连接:在上述树脂中,加入适当DMF后,再加入4.38mmol丁二酸到树脂中,再加入8.76mmol DIPEA,鼓氮气反应30分钟。用2倍树脂体积的DMF 洗涤3次树脂(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。 得到Suc-Ala-Ala-Pro-Phe-CTC Resin。 结构如下:
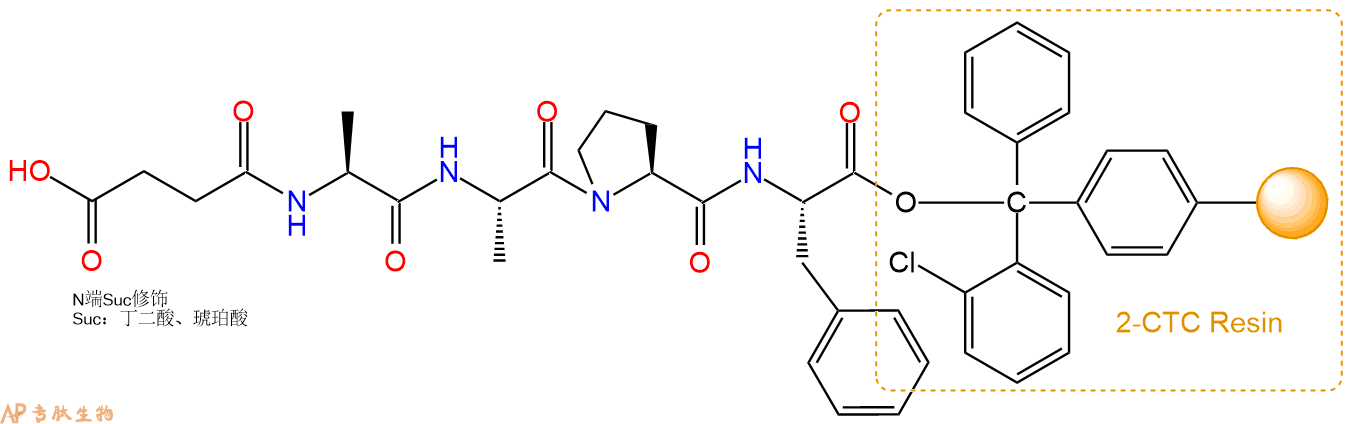
6、全保护切割:配置0.5%TFA/DCM溶液,溶液体积约为树脂体积的3倍。再次用DCM洗涤树脂2遍(去除残留DMF),后将配置好的溶液倒入到反应器中,反应30分钟。抽滤树脂,收集滤液(此时多肽已经从树脂上分离,存在于滤液中)。多肽序列为 Suc-Ala-Ala-Pro-Phe-CTC Resin。 在滤液中添加DIEPA,调PH至7-8。用饱和NaHCO3洗涤滤液,分离出DCM层溶液。可适当旋蒸DCM层溶液,减少有机溶剂。再次加入1或2倍体积的乙酸乙酯,用稀HCl溶液调PH至微酸性,将多肽从DCM层萃取到乙酸乙酯层。用饱和NaCl洗涤2次乙酸乙酯层。用无水硫酸镁吸收乙酸乙酯层的水分。通过减压旋蒸,直接将乙酸乙酯完全旋蒸掉,得到晶体状固体多肽,用于下一步C端反应。或通过减压旋蒸保留适量乙酸乙酯的溶液体积,加入冰乙醚析出 多肽,然后对多肽进行烘干操作即可用于下一步C端反应。Suc-Ala-Ala-Pro-Phe-COOH的结构图如下。
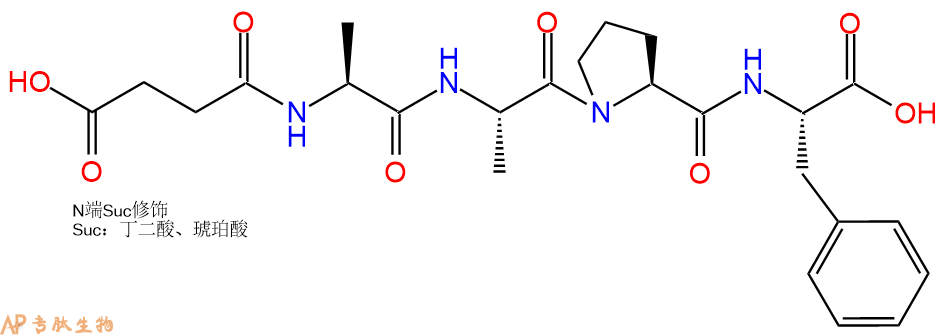
7、4-硝基苯胺反应连接:在上述树脂中,加入适当DMF后,再加入4.38mmol 4-硝基苯胺到树脂中,再加入8.76mmol DIPEA、4.16mmol HBTU,鼓氮气反应30分钟。用2倍树脂体积的DMF 洗涤3次树脂(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。 得到 Suc-Ala-Ala-Pro-Phe-pNA。 结构如下:
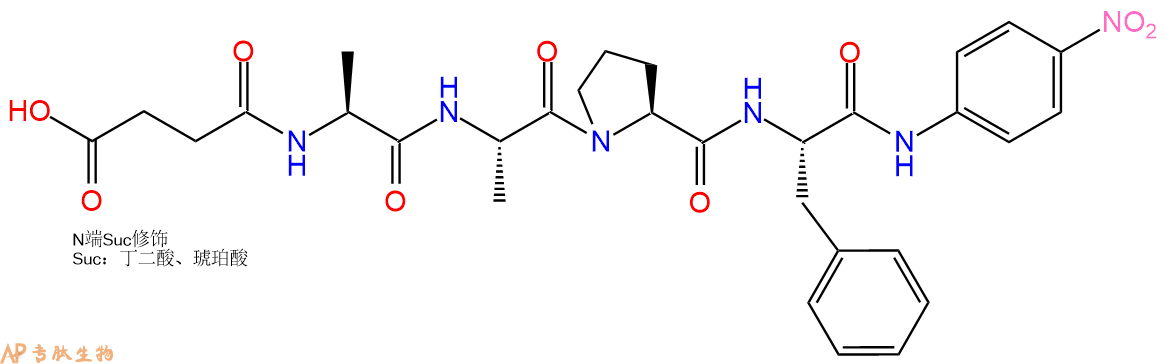
8、切割:6倍树脂体积的切割液(或每1g树脂加8ml左右的切割液),摇床摇晃 2小时,过滤掉树脂,用冰无水乙醚沉淀滤液,并用冰无水乙醚洗涤沉淀物3次,最后将沉淀物放真空干燥釜中,常温干燥24小试,得到粗品Suc-Ala-Ala-Pro-Phe-pNA。结构图见产品结构图。
切割液选择:1)TFA:H2O=95%:5%
2)TFA:H2O:TIS=95%:2.5%:2.5%
3)三氟乙酸:茴香硫醚:1,2-乙二硫醇:苯酚:水=87.5%:5%:2.5%:2.5%:2.5%
(前两种适合没有容易氧化的氨基酸,例如Trp、Cys、Met。第三种适合几乎所有的序列。)
9、纯化冻干:使用液相色谱纯化,收集目标峰液体,进行冻干,获得蓬松的粉末状固体多肽。不过这时要取小样复测下纯度 是否目标纯度。
10、最后总结:
杭州专肽生物技术有限公司(ALLPEPTIDE https://www.allpeptide.com)主营定制多肽合成业务,提供各类长肽,短肽,环肽,提供各类修饰肽,如:荧光标记修饰(CY3、CY5、CY5.5、CY7、FAM、FITC、Rhodamine B、TAMRA等),功能基团修饰肽(叠氮、炔基、DBCO、DOTA、NOTA等),同位素标记肽(N15、C13),订书肽(Stapled Peptide),脂肪酸修饰肽(Pal、Myr、Ste),磷酸化修饰肽(P-Ser、P-Thr、P-Tyr),环肽(酰胺键环肽、一对或者多对二硫键环),生物素标记肽,PEG修饰肽,甲基化修饰肽等。
以上所有内容,为专肽生物原创内容,请勿发布到其他网站上。


















