

400-998-5282
专注多肽 服务科研
400-998-5282
专注多肽 服务科研


多肽具有一个显著的结构性优势,即可以在不影响原有功能性多肽片段的基础上,通过固相合成或生物合成,在多肽的一端或两端引入新的功能性多肽序列,获得多功能性融合肽。 这种具有受体/配体结合能力的pH 敏感脂质体进入细胞的机制,往往不同于普通脂质体的融合方式。如类似RGD/αV β3或者Transferrin /TfR的结合方式,会诱导肿瘤组织细胞主动内吞脂质体继而释放药物/核酸,此类多肽修饰的pH 敏感脂质体药物/核酸释放过程。常见的多肽修饰物按照修饰位点可分为四大类:C 末端修饰(酰胺化、硫酸酯化等)、N 末端修饰(乙酰化、脂肪酸化等)、中间残基修饰(与Se r-、Ty r-、A sn-、Thr-结合的糖基化修饰;与Ser-、Ty r-、Thr-结合的磷酸化修饰等)以及环化修饰。目前对多肽修饰的研究已经成为热点。
化学合成的多肽抗原是小分子, 本身很难具有好的抗原性,只能诱导动物产生很弱的免疫反应,因而与载体蛋白交联是很重要的。载体蛋白含有很多抗原决定基,能够刺激T-帮助细胞,进而诱导 B-细胞反应。用于与多肽交联的载体蛋白有多种,其中常用的是keyhole limpet hemacyanin (KLH), 牛血清白蛋白(bovine serum albumin,BSA), 卵清蛋白(ovalbumin,OVA)和牛甲状腺球蛋白(bovine thyroglobulin,THY)。KLH具有更高的抗原性,是为常用的多肽交联载体。BSA也常用来作为多肽载体,但由于BSA经常被用做检测试验的阻断剂而使得该方法生产的抗体在应用上存在着一定的局限性。
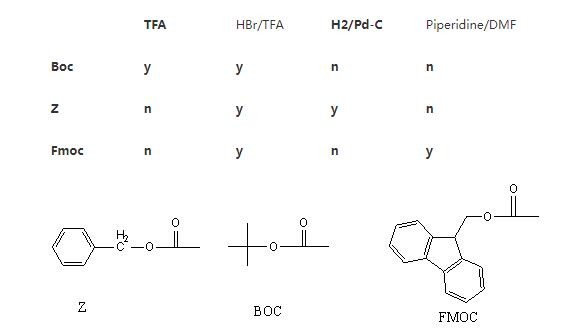
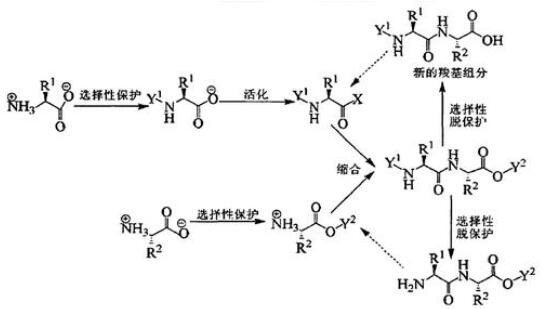
多肽合成(化学)方法,包括液相和固相两种方法。液相多肽合成方法现在主要采用BOC和Z两种保护方法,现在主要应用在短肽合成,如阿斯巴甜,力肽,催产素等,其相对与固相多肽合成,具有保护基选择多,成本低廉,合成规模容易放大的许多优点。与固相多肽合成比较,液相多肽合成主要缺点是,合成范围小,一般都集中在10个氨基酸以内的多肽合成,还有合成中需要对中间体进行提纯,时间长,工作量大。固相多肽合成方法现在主要采用FMOC和BOC两种方法,它具有合成方便,迅速,容易实现自动化,而且可以比较容易的合成到30个氨基酸左右多肽。
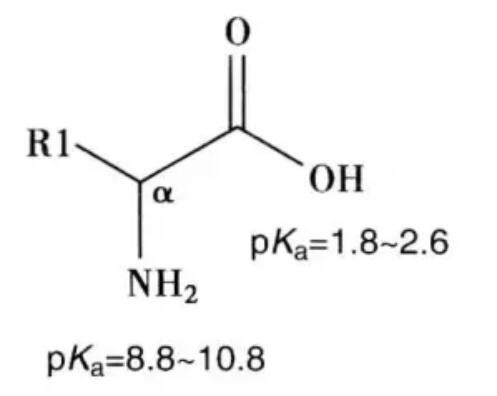
(6)肽单元:参于肽键组成的6个原子(Cα1、C、O、N、H和Cα2)位于同一平面,Cα1和Cα2在平面上所处的位置为反式构型。此同一平面上的6个原子构成钛单元,其中,肽键的键长为0.132nm,介于C-N的单键长(0.149nm)和双链长(0.127nm)之间,所以有一定程度双链性能,不能自由旋转,而Cα分别与N和CO相连的键都是典型的单键,可以自由旋转,正是由于肽单元上Cα原子所连的两个单键可以自由旋转,决定了两个相邻肽单元平面的相对空间位置。
介绍了植物多肽信号的结构和分类,阐述了胞外非分泌型多肽、胞内非分泌型多肽和富含半胱氨酸的分泌型多肽结构特征及生物功能。重点讨论了植物多肽的酪氨酸硫酸化、脯氨酸羟基化和羟脯氨酸阿拉伯糖基化的翻译后修饰方式及蛋白质水解加工方式。并对不同种类翻译后修饰小肽结构和功能进行了概述。

















