

400-998-5282
专注多肽 服务科研
400-998-5282
专注多肽 服务科研
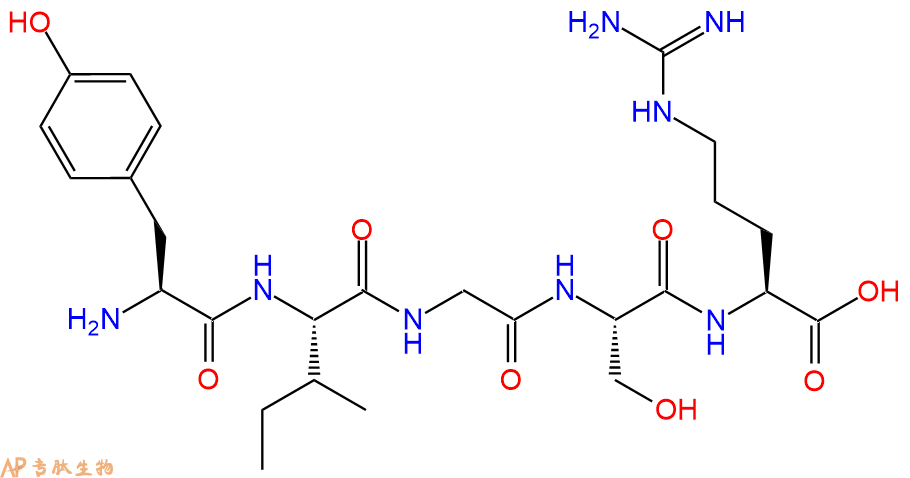
位于层粘连蛋白B1链结构域III内的五肽YIGSR代表层粘连蛋白中的主要受体结合位点。通过与层粘连蛋白竞争细胞表面受体,短肽能够抑制细胞附着在基底膜上,从而防止肿瘤转移的形成。
编号:158449
CAS号:110590-64-2
单字母:H2N-YIGSR-OH
| 编号: | 158449 |
| 中文名称: | Laminin Pentapeptide(929-933)、Laminin (929-933) |
| 英文名: | Laminin Pentapeptide(929-933)、Laminin (929-933) |
| 英文同义词: | Laminin β-1 Chain (929-933) (human, mouse), YIGSR, Laminin Fragment (929-933) |
| CAS号: | 110590-64-2 |
| 单字母: | H2N-YIGSR-OH |
| 三字母: | H2N N端氨基:N-terminal amino group。在肽或多肽链中含有游离a-氨基的氨基酸一端。在表示氨基酸序列时,通常将N端放在肽链的左边。 -TyrL-酪氨酸:tyrosine。系统命名为(2S)-氨基-3-(4-羟基苯基)丙酸。是编码氨基酸。符号:Y,Tyr。 -IleL-异亮氨酸:isoleucine。系统命名为(2S)-氨基-(3R)-甲基戊酸。是编码氨基酸。有两个手性碳原子,是哺乳动物的必需氨基酸。符号:I,Ile。 -Gly甘氨酸:glycine。系统命名为 2-氨基乙酸。是编码氨基酸中没有旋光性的最简单的氨基酸,因具有甜味而得名。符号:G,Gly。 -SerL-丝氨酸:serine。系统命名为(2S)-氨基-3-羟基丙酸。是编码氨基酸。因可从蚕丝中获得而得名。符号:S,Ser。在丝原蛋白及某些抗菌素中含有 D-丝氨酸。 -ArgL-精氨酸:arginine。系统命名为(2S)-氨基-5-胍基戊酸。在生理条件下带正电荷,为编码氨基酸。是幼小哺乳动物的必需氨基酸。符号:R,Arg。 -OHC端羧基:C-terminal carboxyl group。在肽或多肽链中含有游离羧基的氨基酸一端。在表示氨基酸序列时,通常将C端放在肽链的右边。 |
| 氨基酸个数: | 5 |
| 分子式: | C26H42N8O8 |
| 平均分子量: | 594.66 |
| 精确分子量: | 594.31 |
| 等电点(PI): | - |
| pH=7.0时的净电荷数: | 2.97 |
| 平均亲水性: | -0.475 |
| 疏水性值: | -0.42 |
| 外观与性状: | 白色粉末状固体 |
| 消光系数: | 1490 |
| 来源: | 人工化学合成,仅限科学研究使用,不得用于人体。 |
| 纯度: | 95%、98% |
| 盐体系: | 可选TFA、HAc、HCl或其它 |
| 生成周期: | 2-3周 |
| 储存条件: | 负80℃至负20℃ |
| 标签: | 层粘连蛋白(Laminins) |
| 参考文献(References): | G.Hunt, Exp. Cell Biol., 57, 165 (1989) Y.Iwamoto et al., J. Cell. Physiol., 134, 287 (1988) Y.Iwamoto et al., Science, 238, 1132 (1987) J.Graf et al., Biochemistry, 26, 6896 (1987) |
位于层粘连蛋白B1链结构域III内的五肽YIGSR代表层粘连蛋白中的主要受体结合位点。通过与层粘连蛋白竞争细胞表面受体,短肽能够抑制细胞附着在基底膜上,从而防止肿瘤转移的形成。YIGSR还促进肿瘤细胞迁移,此外,还介导多种上皮细胞与层粘连蛋白的粘附。
The pentapeptide YIGSR located within domain III of the laminin B1 chain, represents a major receptor binding site in laminin. By competing with laminin for the cell surface receptor, the short peptide is able to inhibit cell attachment to basement membranes, thus preventing the formation of tumor metastases. YIGSR also promotes tumor cell migration and, moreover, mediates the adhesion of a variety of epithelial cells to laminin.
衍生自层粘连蛋白B1链结构域III的肽代表细胞附着和层粘连蛋白受体结合活性所需的最小序列。它还能抑制注射黑色素瘤细胞的小鼠肺部肿瘤集落的形成。此外,该肽介导多种上皮细胞与层粘连蛋白的粘附。
Peptide derived from domain III of the laminin B1 chain represents minimal sequence necessary for cell attachment and laminin receptor-binding activity. It also inhibits lung tumor colony formation in mice injected with melanoma cells. Moreover, this peptide mediates the adhesion of a variety of epithelical cells to laminin.
层粘连蛋白(929-933)是源自层粘连蛋白的短基序,涉及细胞粘附和整合素结合研究。其序列支持基质相互作用所需最小结构域的评估。研究人员用它探测肽驱动的细胞附着和迁移调节。该肽有助于细胞外基质研究中仿生支架的完善。
Laminin (929-933) is a short laminin-derived motif implicated in cell-adhesion and integrin-binding studies. Its sequence supports evaluation of minimal domains required for matrix interaction. Researchers use it to probe peptide-driven modulation of cell attachment and migration. The peptide aids refinement of biomimetic scaffolds in extracellular matrix research.
Laminins are the protein network that is the foundation for most cells and organs. They are large trimeric proteins, also in the family of glycoproteins that contain an alpha chain, a beta chain and a gamma chain which are found in five, three and three genetic variants. As a family of glycoproteins, laminins are an integral part of the structural scaffolding in almost every tissue of an organism. This makes the laminin a vital part of the maintenance and survival of the tissues. Consequently, defective laminins cause muscular dystrophy, junctional epidermolysis bullosa and defects of the kidney a defect.
Recently, publications have shown that laminins can be used to culture cells that are difficult to culture on other substrates. These trimeric glycoproteins form a similar structure to a cross, giving it a structure that has the ability to bind to other cell membrane and extracellular molecules. The three shorter arms of the laminin are particularly efficient at binding to other laminin molecules, which allows them to form larger sheets. The long arm is capable of binding to cells, which then helps anchor organized tissue cells to the membrane. These laminins are an important and biologically active part of the basal lamina, influenincing cell differentiation, migration and adhesion.
| DOI | 名称 | |
|---|---|---|
| 10.1016/j.biomaterials.2004.07.032 | Immobilization of laminin peptide in molecularly aligned chitosan by covalent bonding | 下载 |
| 10.1111/j.1464-410X.2007.07193.x | Bioluminescence imaging to monitor bladder cancer cell adhesion in vivo: a new approach to optimize a syngeneic, orthotopic, murine bladder cancer model | 下载 |
| 10.1002/bip.20854 | Enhancement of gene transfer using YIGSR analog of Tat-derived peptide | 下载 |
| 10.1159/000163521 | The role of laminin in cancer invasion and metastasis | 下载 |
| 10.1126/science.2961059 | YIGSR, a synthetic laminin pentapeptide, inhibits experimental metastasis formation | 下载 |
| 10.1021/bi00396a004 | A pentapeptide from the laminin B1 chain mediates cell adhesion and binds the 67,000 laminin receptor | 下载 |
| 10.1002/jcp.1041340216 | Synthetic pentapeptide from the B1 chain of laminin promotes B16F10 melanoma cell migration | 下载 |
| 10_1002jbm_a_37460 | Click chemistry functionalization of self-assembling peptide hydrogels | 下载 |
多肽H2N-Tyr-Ile-Gly-Ser-Arg-COOH的合成步骤:
1、合成CTC树脂:称取1.91g CTC Resin(如初始取代度约为0.94mmol/g)和2.15mmol Fmoc-Arg(Pbf)-OH于反应器中,加入适量DCM溶解氨基酸(需要注意,此时CTC树脂体积会增大好几倍,避免DCM溶液过少),再加入5.39mmol DIPEA(Mw:129.1,d:0.740g/ml),反应2-3小时后,可不抽滤溶液,直接加入1ml的HPLC级甲醇,封端半小时。依次用DMF洗涤2次,甲醇洗涤1次,DCM洗涤一次,甲醇洗涤一次,DCM洗涤一次,DMF洗涤2次(这里使用甲醇和DCM交替洗涤,是为了更好地去除其他溶质,有利于后续反应)。得到 Fmoc-Arg(Pbf)-CTC Resin。结构图如下:
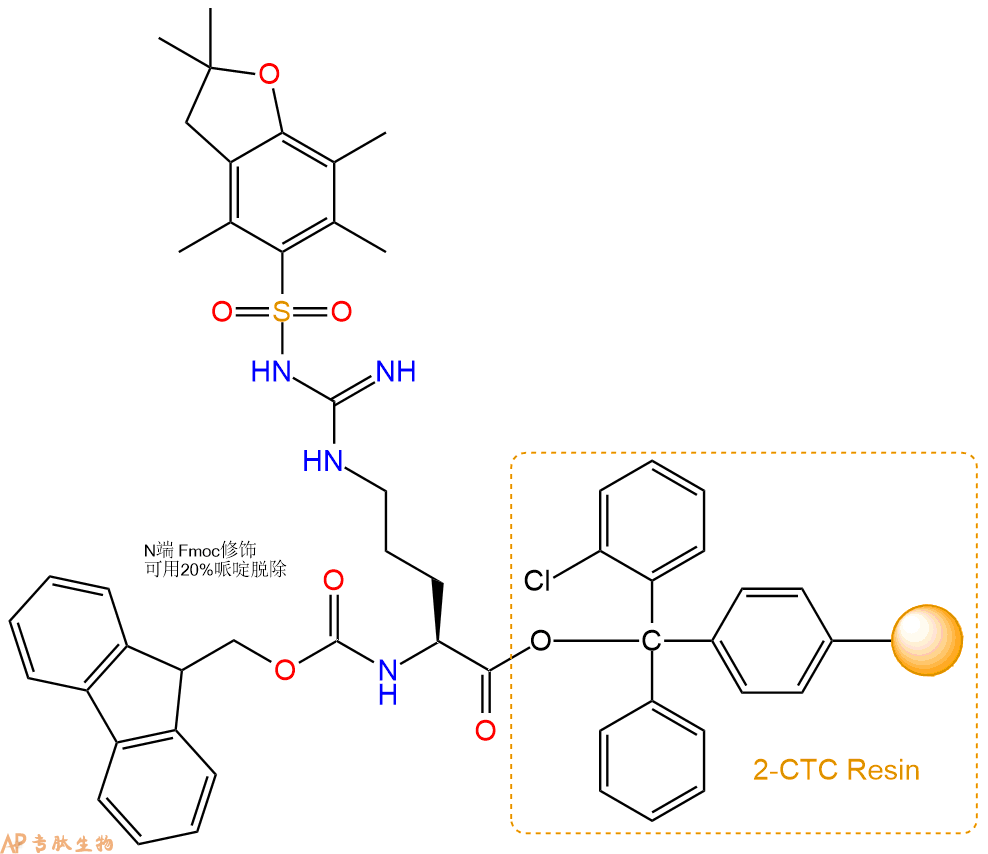
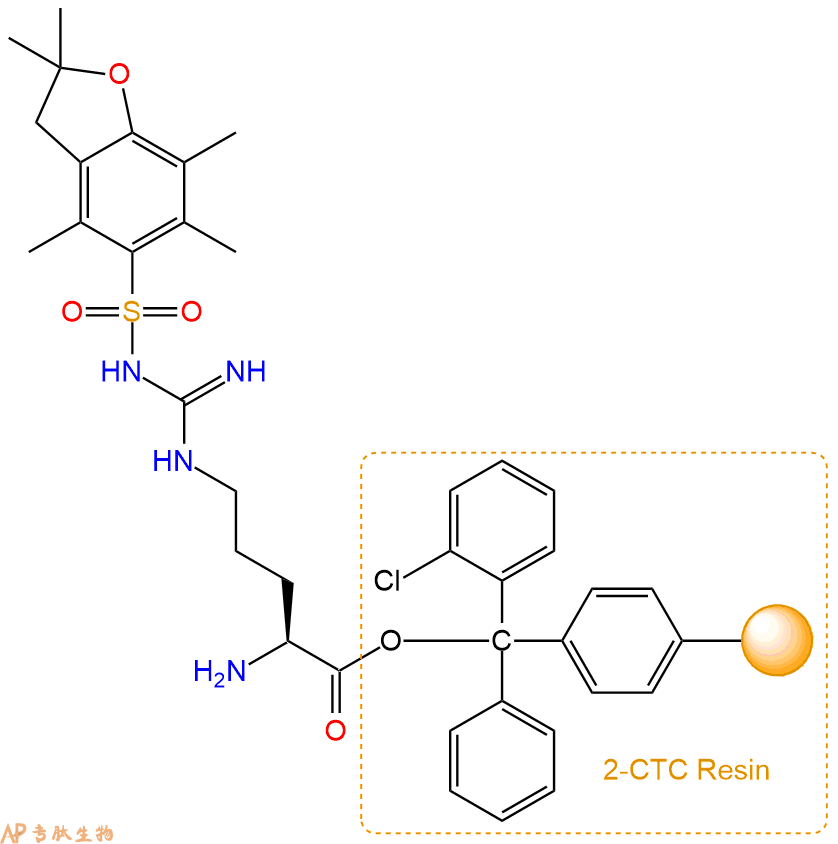
2、脱Fmoc:加3倍树脂体积的20%Pip/DMF溶液,鼓氮气30分钟,然后2倍树脂体积的DMF 洗涤5次。得到 H2N-Arg(Pbf)-CTC Resin 。(此步骤脱除Fmoc基团,茚三酮检测为蓝色,Pip为哌啶)。结构图如下:
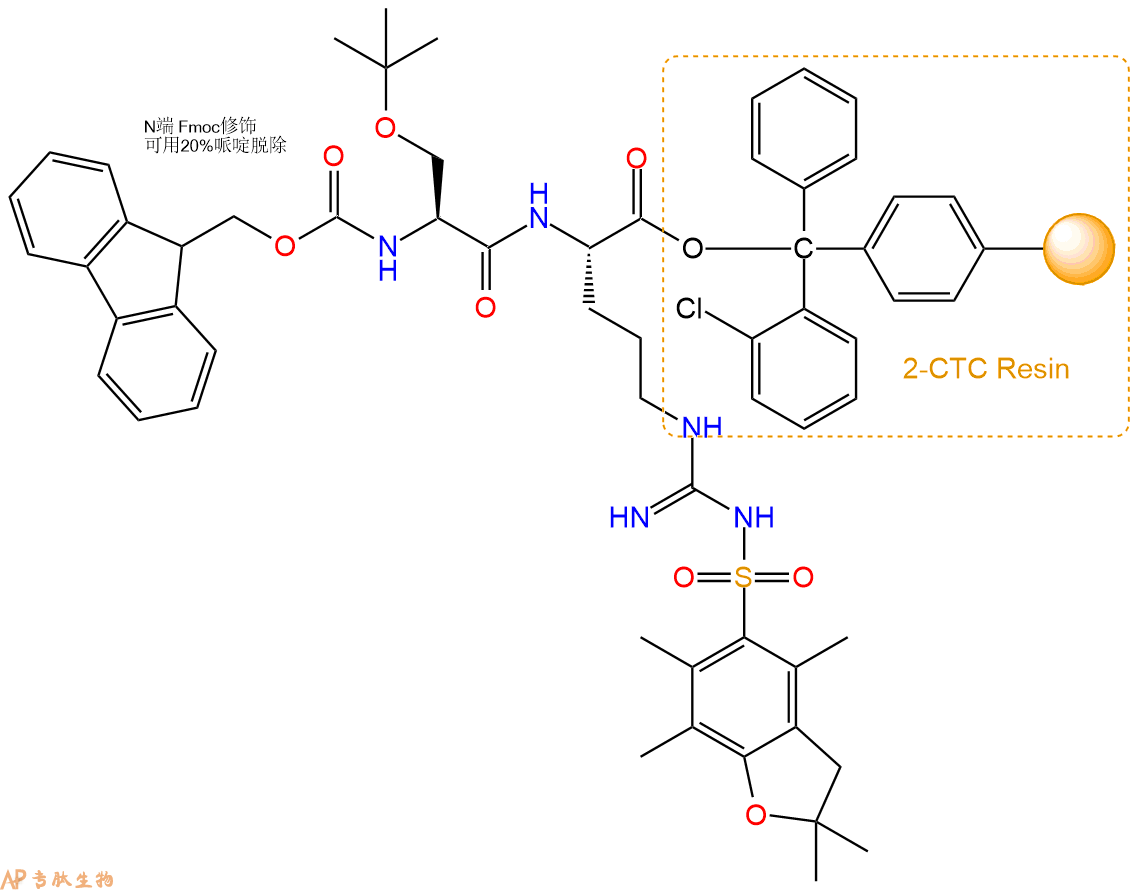
3、缩合:取5.39mmol Fmoc-Ser(tBu)-OH 氨基酸,加入到上述树脂里,加适当DMF溶解氨基酸,再依次加入10.77mmol DIPEA,5.12mmol HBTU。反应30分钟后,取小样洗涤,茚三酮检测为无色。用2倍树脂体积的DMF 洗涤3次树脂。(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。得到Fmoc-Ser(tBu)-Arg(Pbf)-CTC Resin。氨基酸:DIPEA:HBTU:树脂=3:6:2.85:1(摩尔比)。结构图如下:
4、依次循环步骤二、步骤三,依次得到
H2N-Ser(tBu)-Arg(Pbf)-CTC Resin
Fmoc-Gly-Ser(tBu)-Arg(Pbf)-CTC Resin
H2N-Gly-Ser(tBu)-Arg(Pbf)-CTC Resin
Fmoc-Ile-Gly-Ser(tBu)-Arg(Pbf)-CTC Resin
H2N-Ile-Gly-Ser(tBu)-Arg(Pbf)-CTC Resin
Fmoc-Tyr(tBu)-Ile-Gly-Ser(tBu)-Arg(Pbf)-CTC Resin
以上中间结构,均可在专肽生物多肽计算器-多肽结构计算器中,一键画出。
最后再经过步骤二得到 H2N-Tyr(tBu)-Ile-Gly-Ser(tBu)-Arg(Pbf)-CTC Resin,结构如下:
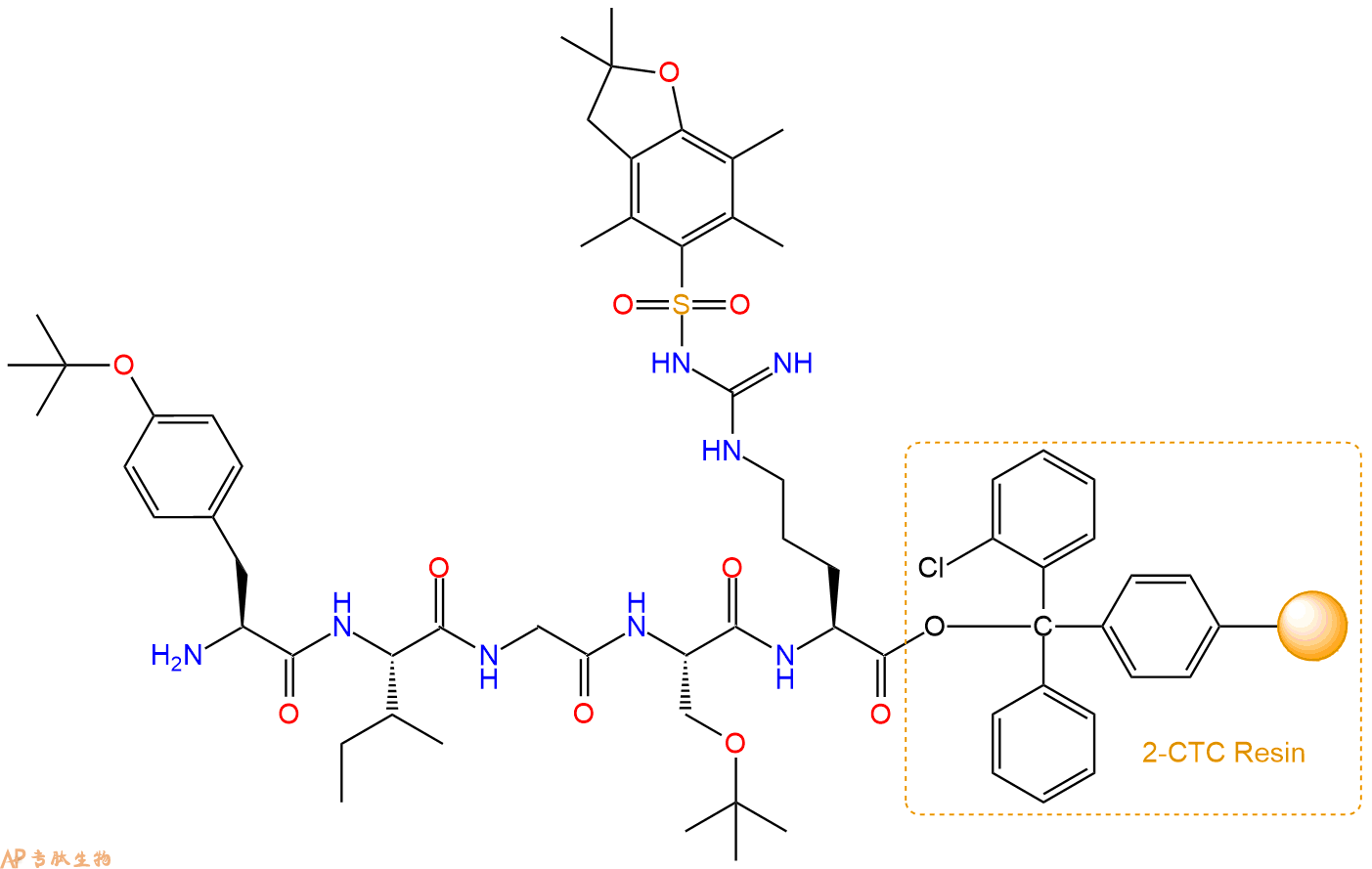
5、切割:6倍树脂体积的切割液(或每1g树脂加8ml左右的切割液),摇床摇晃 2小时,过滤掉树脂,用冰无水乙醚沉淀滤液,并用冰无水乙醚洗涤沉淀物3次,最后将沉淀物放真空干燥釜中,常温干燥24小试,得到粗品H2N-Tyr-Ile-Gly-Ser-Arg-COOH。结构图见产品结构图。
切割液选择:1)TFA:H2O=95%:5%、TFA:H2O=97.5%:2.5%
2)TFA:H2O:TIS=95%:2.5%:2.5%
3)三氟乙酸:茴香硫醚:1,2-乙二硫醇:苯酚:水=87.5%:5%:2.5%:2.5%:2.5%
(前两种适合没有容易氧化的氨基酸,例如Trp、Cys、Met。第三种适合几乎所有的序列。)
6、纯化冻干:使用液相色谱纯化,收集目标峰液体,进行冻干,获得蓬松的粉末状固体多肽。不过这时要取小样复测下纯度 是否目标纯度。
7、最后总结:
杭州专肽生物技术有限公司(ALLPEPTIDE https://www.allpeptide.com)主营定制多肽合成业务,提供各类长肽,短肽,环肽,提供各类修饰肽,如:荧光标记修饰(CY3、CY5、CY5.5、CY7、FAM、FITC、Rhodamine B、TAMRA等),功能基团修饰肽(叠氮、炔基、DBCO、DOTA、NOTA等),同位素标记肽(N15、C13),订书肽(Stapled Peptide),脂肪酸修饰肽(Pal、Myr、Ste),磷酸化修饰肽(P-Ser、P-Thr、P-Tyr),环肽(酰胺键环肽、一对或者多对二硫键环),生物素标记肽,PEG修饰肽,甲基化修饰肽
以上所有内容,为专肽生物原创内容,请勿发布到其他网站上。


















