

400-998-5282
专注多肽 服务科研
400-998-5282
专注多肽 服务科研
甘丙肽是一种肽,属于神经肽家族,存在于内分泌系统,中枢和周围神经系统中。已经证明它可以抑制中枢神经系统中乙酰胆碱的释放
编号:174692
CAS号:114547-31-8
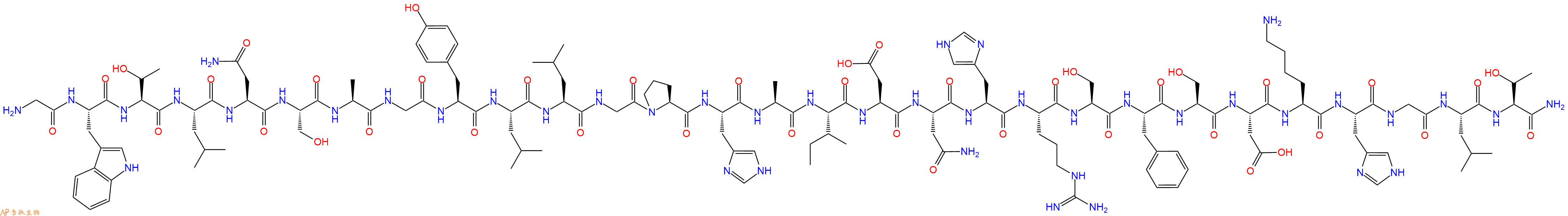
单字母:H2N-GWTLNSAGYLLGPHAIDNHRSFSDKHGLT-CONH2
| 参考文献(References): | M.E. Vrontakis et al., JBC, 262, 16755 (1987) L.M. Kaplan et al., PNAS, 85, 1065 (1988) |
Galanin (1-29)(rat, mouse) TFA 是一种非选择性甘丙肽受体 (galanin receptor) 激动剂,对 GAL1、GAL2 和 GAL3 的 Ki 值分别为 0.98、1.48 和 1.47 nM。具有抗惊厥作用。
Galanin (1-29)(rat, mouse) TFA is a non-selective galanin receptor agonist, with Kis of 0.98, 1.48 and 1.47 nM for GAL1, GAL2 and GAL3, respectively. Anticonvulsant effect.
甘丙肽是一种肽,属于神经肽家族,存在于内分泌系统,中枢和周围神经系统中。已经证明它可以抑制中枢神经系统中乙酰胆碱的释放,有趣的是,在阿尔茨海默病患者中,已经发现甘丙肽纤维的过度矿化。甘丙肽还具有多种生理作用,包括抑制胰岛素释放,刺激催乳素和生长激素分泌,并在进食,情绪,认知,神经内分泌调节中发挥作用,并影响胃肠动力。甘丙肽通过与G蛋白偶联受体相关而发挥作用。
Galanin is a peptide that belongs to the family of neuropeptides and is found in the endocrine system, the central and peripheral nervous systems. It has been shown to inhibit the release of acetylcholine in the central nervous system and interestingly in Alzheimer’s disease patients, hyperinervation of galanin fibres has been found. Galanin is also known to have a variety of physiological effects including inhibition of insulin release, stimulation of prolactin and growth hormone secretion and it also plays a role in feeding, mood, cognition, neuroendocrine regulation and affects gastrointestinal motility. Galanin asserts its effects through associated with G-protein coupled receptors.
大鼠源甘丙肽是一种保守神经肽,其 N 端疏水核心可稳定部分螺旋基序。其序列有助于受体识别决定因素定位。研究者通过评估构象来探究结构‑功能关系。该分子有助于神经内分泌多肽研究。
Galanin, rat, is a conserved neuropeptide with an N-terminal hydrophobic core that stabilizes partial helical motifs. Its sequence supports mapping of receptor-recognition determinants. Researchers evaluate conformation to explore structure-function relationships. The molecule aids neuroendocrine peptide studies.
Galanin (mouse, rat) 是一种非选择性甘丙肽受体 (galanin receptor) 激动剂,对 GAL1、GAL2 和 GAL3 的 Ki 值分别为 0.98、1.48 和 1.47 nM。具有抗惊厥作用。
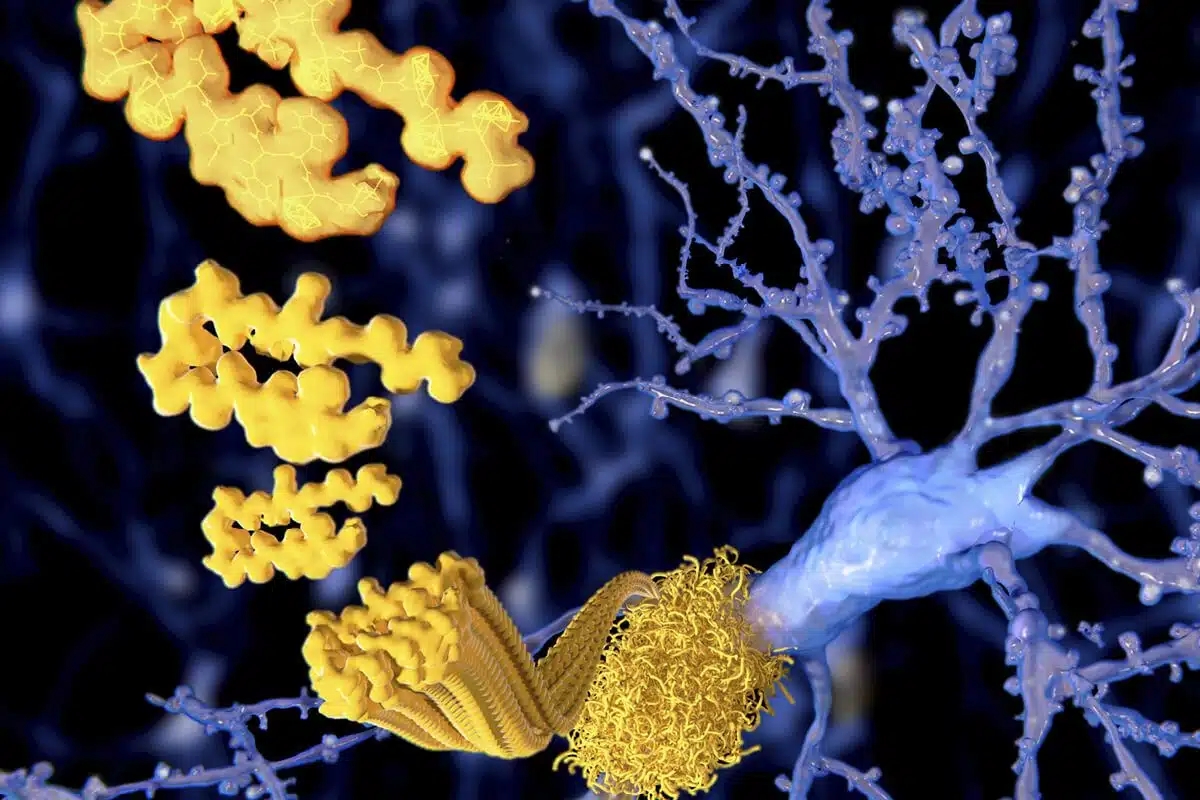
Extracellular amyloid-β peptide deposition into cerebellar plaques and formation of intracellular neurofibrillary fibers accompanied by the loss of neurons are characteristic histopathological lesions found in the brains of Alzheimer‘s disease patients. Individuals suffering from this disease show a gradual loss of cognitive functions and disturbances in behavior. Apart from some rare familial forms of the disease, the onset of Alzheimer‘s disease is usually above 60 years. Since the risk to develop the disease increases with age, Alzheimer‘s disease has turned into a major health and social problem in “first world” countries with an increasing proportion of older people, and is going to become one in emerging states. In this brochure we present amyloid peptides and related products for Alzheimer‘s disease research.
ALZHEIMER’S DISEASE
Alzheimer‘s disease (AD) is the prevalent cause of dementia in elderly people and has become one of the leading causes of death in developed countries together with cardiovascular disorders, cancer, and stroke. It is estimated that more than 46 millions of people suffer from AD all over the world. As age advances, the risk for developing AD increases. The frequency of AD at the age of 60-64 is about 1% and doubles approximately every five years. By the age of 90 and older, approximately 50% of the population suffers from this disease. AD is an irreversible and progressive neurodegenerative disorder. Symptoms include gradual loss of cognitive functions such as memory, verbal and visuospatial abilities, changes in personality, behavior, and activities of daily living. AD patients in the final stages are completely dependent on the care of others.
The characteristic lesions in the brains of AD patients were first described by the German neuropsychiatrist Alois Alzheimer in 1906 during the post-mortem examination of a mentally ill patient whose deterioration he had observed until her death. The lesions consisted of dense extracellular deposits, now designated as neuritic or senile plaques, and intracellular dense bundles of fibrils, which are now known as neurofibrillary tangles.
Currently, diagnosis of AD with adequate testing is approximately 90% accurate. It is based on the exclusion of a variety of diseases causing similar symptoms and a careful neurological and psychiatric examination, as well as neuropsychological testing. Imaging technologies for detecting amyloid plaques and tangles in vivo are becoming more precise and thus a valuable additional tool. Numerous potential biomarkers as α1 -antitrypsin, complement factor H, α2 -macroglobulin, apolipoprotein J, and apolipoprotein A-I for diagnosing AD are being evaluated. However, post-mortem histopathological examination of the brain is still the only definite diagnosis of this disease.
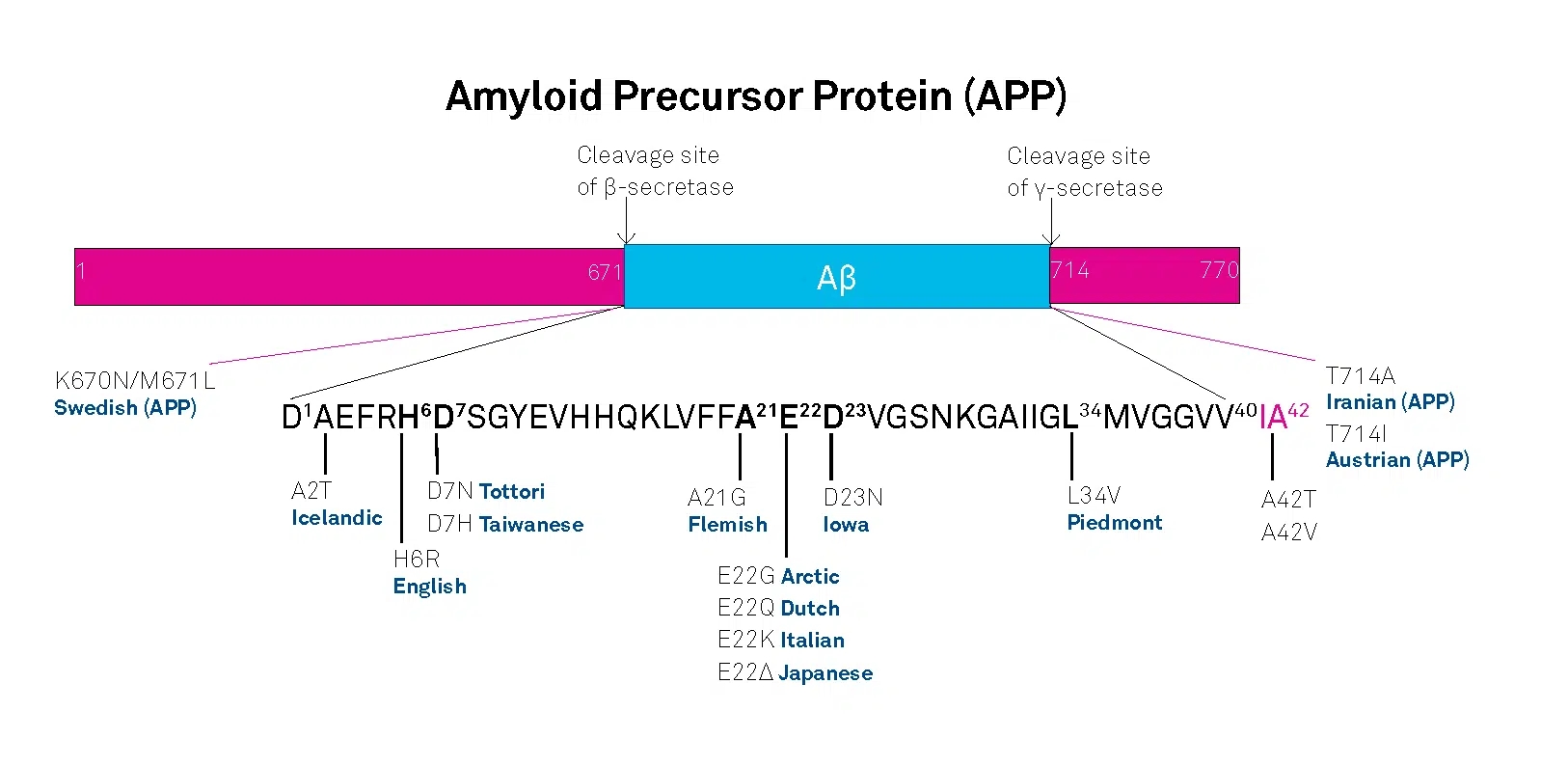
AD can be either inherited or sporadic. The inherited or familial AD is rare and comprises only 5-10% of all cases. Autosomal dominant mutations in the amyloid β/A4 protein precursor (APP) gene on chromosome 21 and the presenilin-1 or -2 genes on chromosomes 14 and 1, respectively, have been attributed to the early onset (before the age of 65) of this disease.
APP belongs to the type-1 integral membrane glycoproteins with at least 10 isoforms generated by alternative splicing of the 19 exons. The predominant transcripts are APP695, APP751, and APP770. A number of mutations within the APP gene have been detected in families with an inherited risk for early onset of AD. Usually, they are named after the region, in which they have been detected, e.g. the London APP717 mutations (V717I, V717F, V717G), the Swedish APP670/671 double mutation (K670N/M671L), the Flemish APP692 mutation (A692G), or the Dutch APP693 mutation (E693Q). The Swedish mutation of the β-secretase cleavage site of APP and mutations of positions 692-694 (Aβ 21-23), which strongly influence the aggregation behavior of Aβ, have been studied intensively.
A choice of relevant mutations in the Aβ region of APP is assembled in the table below.
| Exchanged Position in APP | Exchanged Position in Aβ | Designation |
|---|---|---|
| A673T | A2T | Icelandic |
| H677R | H6R | English |
| D678H | D7H | Taiwanese |
| D678N | D7N | Tottori |
| A692G | A21G | Flemish |
| E693D | E22∆ | Osaka |
| E693G | E22G | Arctic |
| E693Q | E22Q | Dutch |
| E693K | E22K | Italian |
| D694N | D23N | Iowa |
| L705V | L34V | Piedmont |
The presenilins are another group of proteins involved in the development of AD. Presenilins are integral membrane proteins with eight transmembrane domains localized in the endoplasmic reticulum and the Golgi apparatus. A multitude of mutations within the presenilin-1 and two within the presenilin-2 gene account for most of the cases of early onset of AD.
Genetic factors may contribute as well to the late onset of AD. Increased susceptibility is associated with the expression of different apolipoprotein E (ApoE) isoforms due to the polymorphism in the APOE gene on chromosome 19. In the central nervous system, ApoE has been implicated in growth and repair during development or after injury. Carriers of the APOEε4 allele show a higher risk in developing the disease than carriers of the other two possible alleles APOEε2 and APOEε3. The ApoEε4 effect seems to be dose-dependent since individuals with two of these alleles seem to be at two-fold higher risk to develop the disease than those with one allele. Polymorphisms of the α2 -macroglobulin gene on chromosome 12 and the gene coding low-density lipoprotein receptor-related protein 1 (LRP1), LRP1-C/T, have also been suggested to be a risk factor for the late onset of AD. However, further studies in this field are required.
A number of additional, most diverse risk factors have been proposed. These include gender, ethnic group, head trauma, cardiovascular diseases, and educational level.
AD THERAPEUTIC STRATEGIES RELY ON DETAILED KNOWLEDGE OF THE MOLECULES INVOLVED
Women, Hispanics, individuals who have experienced a head trauma earlier in life, and persons who suffer from cardiovascular diseases appear to have a higher risk of developing the disease.
The etiology of AD is still not completely understood. Initial research focused upon determining the molecular structure of the senile plaques and the neurofibrillary tangles originally described by Alois Alzheimer. The main constituents of the senile plaques were identified as cleavage products of APP, designated as amyloid β-peptides (Aβ peptides).
Depending on the composition and the fraction of fibrillar to non-fibrillar forms of these amyloid peptides, several kinds of senile plaques can be distinguished. Three types of proteases, α-secretase, β-secretase (or β-site APP-cleaving enzyme, BACE), and γ-secretase are involved in APP processing. APP can either be processed by the α- and γ- or by the β- and γ-secretases. The major two amyloid peptides identified in senile plaques, amyloid β-protein (1-40) (Aβ40) and amyloid β-protein (1-42) (Aβ42), are generated by successive proteolysis of APP by β- and γ-secretases. Cleavage of APP by β-secretase results in the release of the extracellular N-terminal protein fragment known as soluble APP-β molecule (sAPP-β). Then, the membrane-retained APP is further processed within the transmembrane domain by γ-secretase to yield either Aβ40 or Aβ42. The formation of Aβ40 and Aβ42 is a normal process, and both peptides can be detected in the plasma and cerebrospinal fluid (CSF) of healthy subjects.
In most studies, similar concentrations of Aβ40 have been measured in the CSF of both healthy controls and AD patients. On the other hand, Aβ42 concentrations in the CSF of AD patients are significantly lower than in normal controls, probably reflecting an increased deposition as insoluble plaques.
The neurofibrillary tangles found inside neurons of Alzheimer’s brains are composed of paired helical filaments whose main components are hyperphosphorylated forms of tau, a microtubule associated protein involved in promoting microtubule assembly and stabilization. Self-assembly into paired helical filaments is believed to be a result of hyperphosphorylation due to either the increased activity of protein kinases or the decreased activity of phosphatases.
Several lines of evidence support the view that the accumulation of Aβ42 in the brain is a primary event in the development of AD. Increased cerebral Aβ production appears to be characteristic for all the mutations within the APP and the presenilin genes of familial AD. In patients with Down syndrome (trisomy 21), elevated levels of APP and Aβ due to a third copy of the APP gene result in deposition of Aβ at an early age between 20 and 30.
Formation of neurofibrillary tangles is considered as a consequence of Aβ deposition with a further impact on the progression of the disease possibly due to disruption of axonal transport mechanisms in neurons.
The detailed knowledge about the molecules involved in AD has led to the development of several therapeutic strategies.
One strategy aims at the reduction of Aβ40 and Aβ42 by inhibition of either β- or γ-secretase activity or by clearance of Aβ in the brain by means of immunization with these peptides. Transition metals as Cu, Fe and Zn play an important role in the pathology of AD. Aggregation and neurotoxicity of Aβ are dependent on the presence of copper, so Cu-chelating agents showed promising effects in animal models. Another approach is the prevention of the cellular inflammatory response in the cerebral cortex elicited by the progressive accumulation of Aβ. Further preventive therapeutic strategies are based on the findings that cholesterol-lowering drugs such as statins and estrogen replacement therapy reduce the risk of developing AD. An additional treatment alternative would be the inhibition of the serine-threonine protein kinases, glycogen synthase kinase 3 (GSK3) and cyclin-dependent kinase 5 (CDK5), which are probably responsible for the phosphorylation of the tau protein. Inhibition of calpain, an enzyme showing increased activity in AD brains, led to promising results in animal studies. Calpain cleaves the CDK5 activator p35 leading to p25 formation and CDK5 overactivation.
Several acetylcholinesterase inhibitors such as tacrine, donepezil, rivastigmine, and galantamine have been approved for the treatment of mild to moderate AD by the FDA and other authorities. They act by reducing the deficits of the neurotransmitter acetylcholine associated with cognitive impairment in AD patients. The amantadine derivative memantine, an NMDA receptor antagonist, which was already used for the treatment of moderate to severe AD in Europe, has gained approval in the United States by the FDA as well.
A promising drug candidate, the β-secretase inhibitor verubecestat (MK-8931) developed for the management of mild to moderate AD, has moved to phase III. Moreover, the BACE inhibitor AZD3293 showed encouraging results in clinical studies. Antibodies as aducanumab and solanezumab, which have been designed to degrade plaques and lower the level of Aβ in the brain, have reached advanced stages of clinical testing for mild cases of AD.
Despite the many promising therapeutic approaches, AD still remains a major burden for the patients, their relatives, and the society.
Wang S, et al. Cloning and expressional characterization of a novel galanin receptor. Identification of different pharmacophores within galanin for the three galanin receptor subtypes. J Biol Chem. 1997;272(51):31949-31952. : https://pubmed.ncbi.nlm.nih.gov/9405385/
Mazarati A, et al. Regulation of kindling epileptogenesis by hippocampal galanin type 1 and type 2 receptors: The effects of subtype-selective agonists and the role of G-protein-mediated signaling. J Pharmacol Exp Ther. 2006;318(2):700-708. : https://pubmed.ncbi.nlm.nih.gov/16699066/
多肽H2N-Gly-Trp-Thr-Leu-Asn-Ser-Ala-Gly-Tyr-Leu-Leu-Gly-Pro-His-Ala-Ile-Asp-Asn-His-Arg-Ser-Phe-Ser-Asp-Lys-His-Gly-Leu-Thr-NH2的合成步骤:
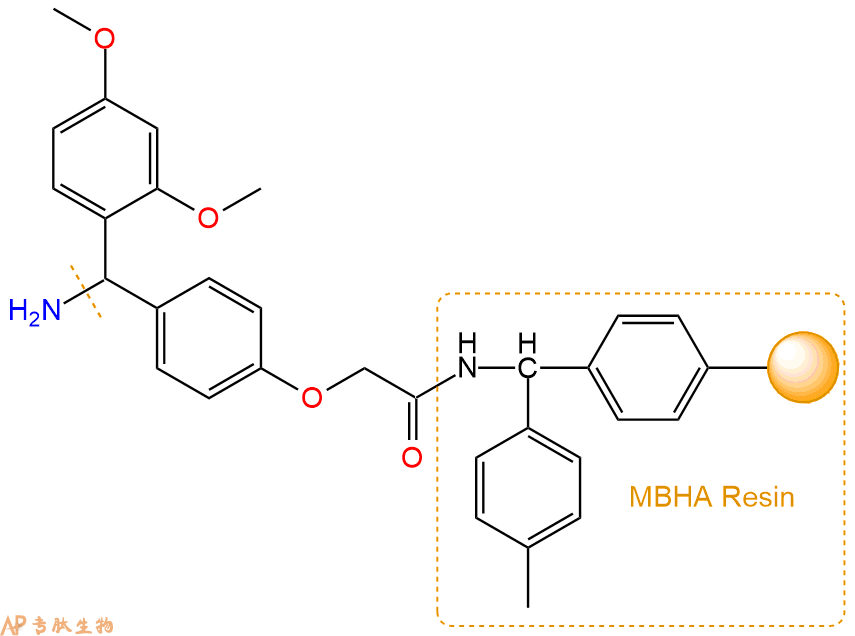
1、合成MBHA树脂:取若干克的MBHA树脂(如初始取代度为0.5mmol/g)和1倍树脂摩尔量的Fmoc-Linker-OH加入到反应器中,加入DMF,搅拌使氨基酸完全溶解。再加入树脂2倍量的DIEPA,搅拌混合均匀。再加入树脂0.95倍量的HBTU,搅拌混合均匀。反应3-4小时后,用DMF洗涤3次。用2倍树脂体积的10%乙酸酐/DMF 进行封端30分钟。然后再用DMF洗涤3次,甲醇洗涤2次,DCM洗涤2次,再用甲醇洗涤2次。真空干燥12小时以上,得到干燥的树脂{Fmoc-Linker-MHBA Resin},测定取代度。这里测得取代度为 0.3mmol/g。结构如下图:
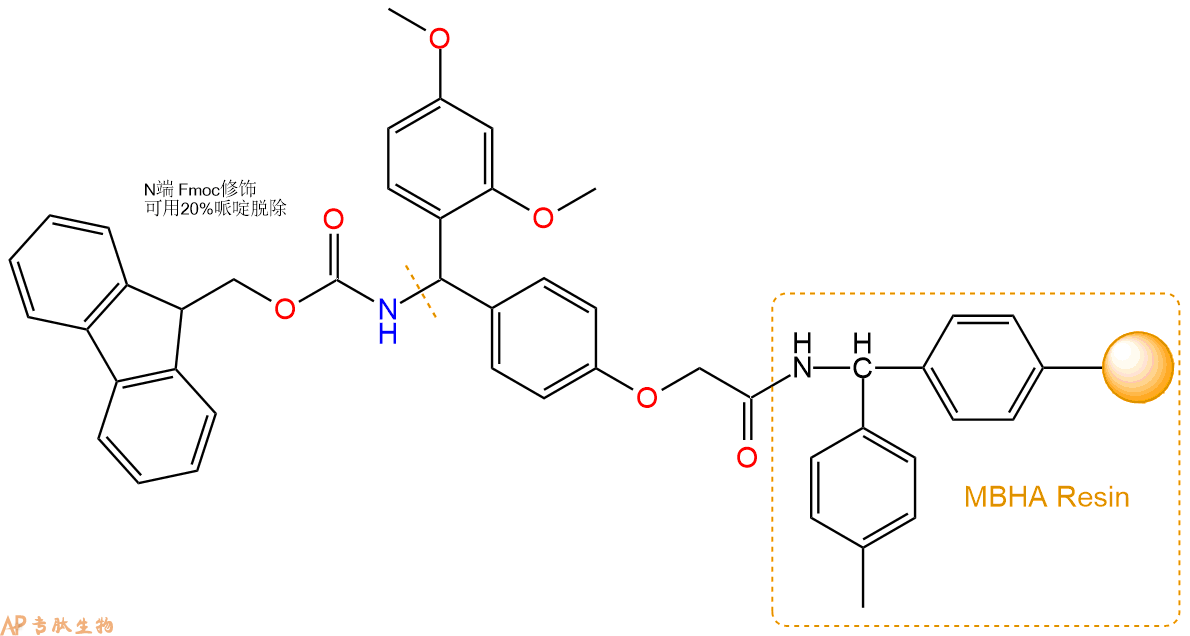
2、脱Fmoc:取1.14g的上述树脂,用DCM或DMF溶胀20分钟。用DMF洗涤2遍。加3倍树脂体积的20%Pip/DMF溶液,鼓氮气30分钟,然后2倍树脂体积的DMF 洗涤5次。得到 H2N-Linker-MBHA Resin 。(此步骤脱除Fmoc基团,茚三酮检测为蓝色,Pip为哌啶)。结构图如下:
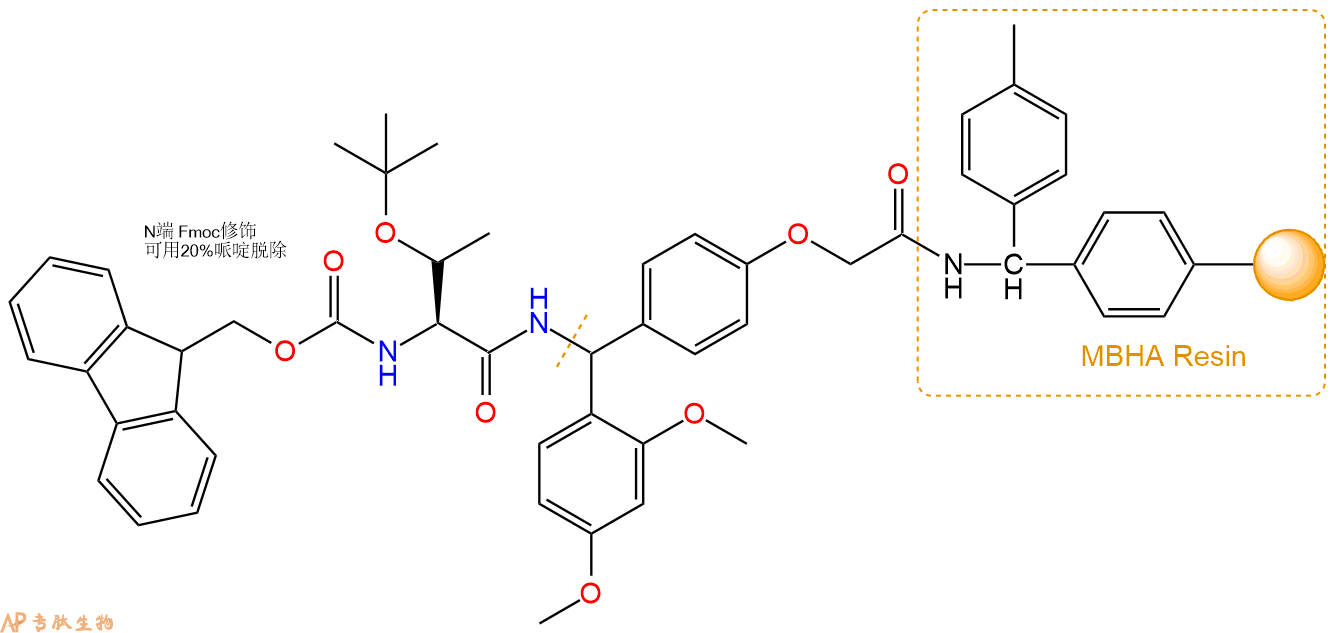
3、缩合:取1.03mmol Fmoc-Thr(tBu)-OH 氨基酸,加入到上述树脂里,加适当DMF溶解氨基酸,再依次加入2.05mmol DIPEA,0.97mmol HBTU。反应30分钟后,取小样洗涤,茚三酮检测为无色。用2倍树脂体积的DMF 洗涤3次树脂。(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。得到Fmoc-Thr(tBu)-Linker-MBHA Resin。氨基酸:DIPEA:HBTU:树脂=3:6:2.85:1(摩尔比)。结构图如下:
4、依次循环步骤二、步骤三,依次得到
H2N-Thr(tBu)-Linker-MBHA Resin
Fmoc-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Leu-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Leu-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Thr(tBu)-Leu-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Thr(tBu)-Leu-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Trp(Boc)-Thr(tBu)-Leu-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
H2N-Trp(Boc)-Thr(tBu)-Leu-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
Fmoc-Gly-Trp(Boc)-Thr(tBu)-Leu-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin
以上中间结构,均可在专肽生物多肽计算器-多肽结构计算器中,一键画出。
最后再经过步骤二得到 H2N-Gly-Trp(Boc)-Thr(tBu)-Leu-Asn(Trt)-Ser(tBu)-Ala-Gly-Tyr(tBu)-Leu-Leu-Gly-Pro-His(Trt)-Ala-Ile-Asp(OtBu)-Asn(Trt)-His(Trt)-Arg(Pbf)-Ser(tBu)-Phe-Ser(tBu)-Asp(OtBu)-Lys(Boc)-His(Trt)-Gly-Leu-Thr(tBu)-Linker-MBHA Resin,结构如下:
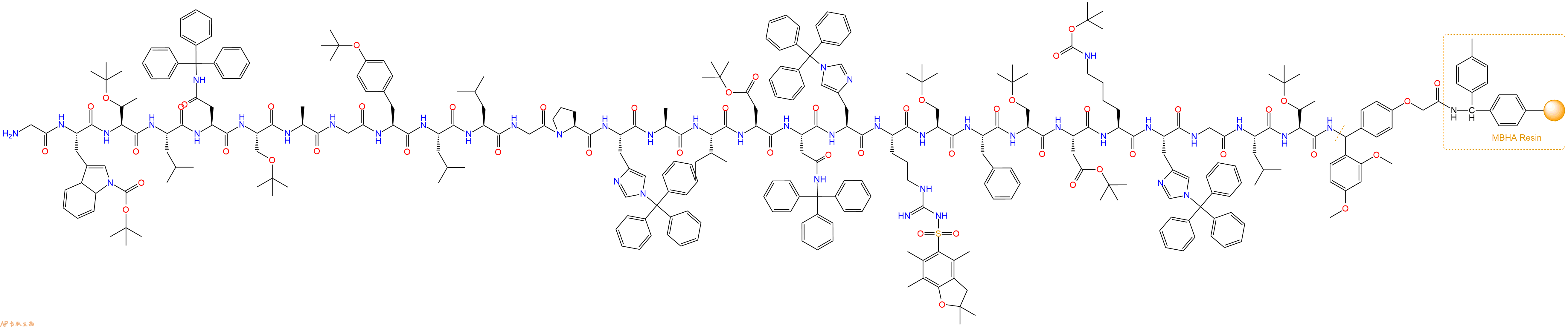
5、切割:6倍树脂体积的切割液(或每1g树脂加8ml左右的切割液),摇床摇晃 2小时,过滤掉树脂,用冰无水乙醚沉淀滤液,并用冰无水乙醚洗涤沉淀物3次,最后将沉淀物放真空干燥釜中,常温干燥24小试,得到粗品H2N-Gly-Trp-Thr-Leu-Asn-Ser-Ala-Gly-Tyr-Leu-Leu-Gly-Pro-His-Ala-Ile-Asp-Asn-His-Arg-Ser-Phe-Ser-Asp-Lys-His-Gly-Leu-Thr-NH2。结构图见产品结构图。
切割液选择:1)TFA:H2O=95%:5%
2)TFA:H2O:TIS=95%:2.5%:2.5%
3)三氟乙酸:茴香硫醚:1,2-乙二硫醇:苯酚:水=87.5%:5%:2.5%:2.5%:2.5%
(前两种适合没有容易氧化的氨基酸,例如Trp、Cys、Met。第三种适合几乎所有的序列。)
6、纯化冻干:使用液相色谱纯化,收集目标峰液体,进行冻干,获得蓬松的粉末状固体多肽。不过这时要取小样复测下纯度 是否目标纯度。
7、最后总结:
杭州专肽生物技术有限公司(ALLPEPTIDE https://www.allpeptide.com)主营定制多肽合成业务,提供各类长肽,短肽,环肽,提供各类修饰肽,如:荧光标记修饰(CY3、CY5、CY5.5、CY7、FAM、FITC、Rhodamine B、TAMRA等),功能基团修饰肽(叠氮、炔基、DBCO、DOTA、NOTA等),同位素标记肽(N15、C13),订书肽(Stapled Peptide),脂肪酸修饰肽(Pal、Myr、Ste),磷酸化修饰肽(P-Ser、P-Thr、P-Tyr),环肽(酰胺键环肽、一对或者多对二硫键环),生物素标记肽,PEG修饰肽,甲基化修饰肽等。
以上所有内容,为专肽生物原创内容,请勿发布到其他网站上。


















