

400-998-5282
专注多肽 服务科研
400-998-5282
专注多肽 服务科研
编号:168678
CAS号:1802086-28-7
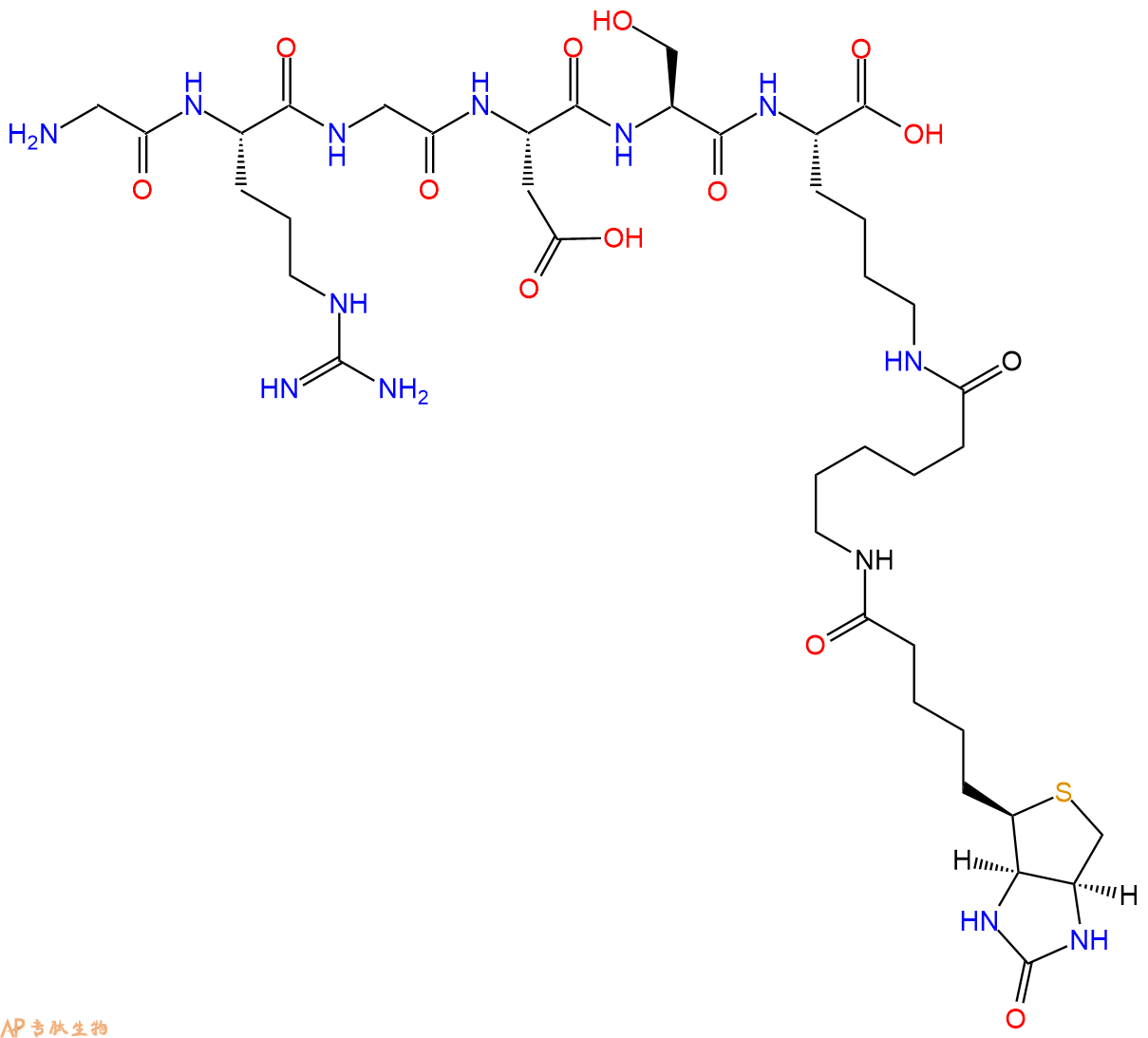
单字母:Biotinyl-HSQGTFTSDYSKYLDSRRAQDFVQWLMNT-OH
胰高血糖素(Glucagon)的定义
胰高血糖素是众所周知的一种胰岛分泌的肽激素,它参与葡萄糖代谢的控制。
Glucagon is best known as a peptide hormone secreted from pancreatic islets that participates in control of glucose metabolism.
胰高血糖素(Glucagon)的相关肽
胰高血糖素被合成为胰高血糖素,并经蛋白水解加工以在胰岛的α细胞内产生胰高血糖素。胰高血糖素原也在肠内表达,在那里它不被加工成胰高血糖素,而是被加工成胰高血糖素样肽家族。在胰腺和肠道中,都会产生三种类型的产品:
Glucagon is synthesized as proglucagon and proteolytically processed to yield glucagons within alpha cells of the pancreatic islets. Proglucagon is also expressed within the intestinal tract, where it is processed not into glucagon, but to a family of glucagon-like peptides. In both pancreas and gut, three types of products are generated:
1、具有已知生物学活性的肽:胰高血糖素和胰高血糖素样肽1(GLP-1)
Peptides with known biological activity: glucagon and glucagon-like peptide-1 (GLP-1)
2、可能具有生物学活性,但表征不佳或仅在非生理浓度下才具有活性的肽:胰高血糖素样肽2(GLP-2)和胃泌酸调节素
Peptides that may have biological activity, but which are poorly characterized or active only at what are considered non-physiologic concentrations: glucagon-like peptide-2 (GLP-2) and oxyntomodulin
3、没有明显生物学活性的肽:glicentin,与glicentin相关的胰肽,主要胰高血糖素原片段【1】。
Peptides without apparent biological activity: glicentin, glicentin-related pancreatic peptide, major proglucagon fragment【1】.
胰高血糖素(Glucagon)的发现
胰高血糖素最初由罗切斯特大学的CP Kimball和John R. Murlin于1923年纯化。胰高血糖素的氨基酸序列在1950年代后期进行了描述【2】。
Glucagon was initially purified by C. P. Kimball and John R. Murlin of the University of Rochester in 1923. The amino acid sequence of glucagon was described in the late-1950s2.
胰高血糖素(Glucagon)的结构特征
胰高血糖素是由29个氨基酸组成的线性肽。它的主要序列在脊椎动物中几乎是完全保守的,并且在结构上与肽激素1的促分泌素家族有关【1】。
Glucagon is a linear peptide of 29 amino acids. Its primary sequence is almost perfectly conserved among vertebrates, and it is structurally related to the secretin family of peptide hormones【1】.
胰高血糖素(Glucagon)的行动方式
胰高血糖素通过其在细胞表面的受体发出信号。胰高血糖素 与胰高血糖素受体的细胞外环的结合导致 后者的构象变化,从而导致随后的偶联的G蛋白的活化。 已知至少两类G蛋白 与胰高血糖素受体的信号转导有关并参与其中, 即G s 和G q。G s 的激活导致 腺苷酸环化酶的激活,细胞内cAMP水平的增加,以及 随后蛋白激酶A(PKA)的激活。激活 的G q导致磷脂酶C,生产的激活 肌醇1,4,5-三磷酸,并随后释放细胞内 钙【3】。
Glucagon signals through its receptor on the cell surface. The binding of glucagon to the extracellular loops of the glucagon receptor results in conformational changes of the latter, leading to subsequent activation of the coupled G proteins. At least two classes of G proteins are known to be associated with and involved in the signal transduction of the glucagon receptor, namely Gs and Gq. The activation of Gs leads to activation of adenylate cyclase, increase in intracellular cAMP levels, and subsequent activation of protein kinase A (PKA). The activation of Gq leads to the activation of phospholipase C, production of inositol 1,4,5-triphosphate, and subsequent release of intracellular calcium【3】.
胰高血糖素(Glucagon)的功能
胰高血糖素在葡萄糖代谢和体内平衡中起关键作用。通过增加糖异生和减少糖酵解来调节血糖。胰岛素的反调节激素可响应胰岛素诱导的低血糖而升高血浆葡萄糖水平。在引发和维持糖尿病的高血糖状况中起重要作用。
Glucagon plays a key role in glucose metabolism and homeostasis. Regulates blood glucose by increasing gluconeogenesis and decreasing glycolysis. A counterregulatory hormone of insulin, raises plasma glucose levels in response to insulin-induced hypoglycemia. Plays an important role in initiating and maintaining hyperglycemic conditions in diabetes.
GLP-1是葡萄糖依赖性胰岛素释放的有效刺激剂。在胃动力和血浆胰高血糖素水平的抑制中起重要作用。可能与胰岛素的作用无关地抑制饱腹感并刺激周围组织中的葡萄糖释放。对肠上皮具有促进生长的活性。还可能通过对LH,TSH,CRH,催产素和加压素分泌的影响来调节下丘脑垂体轴(HPA)。通过刺激胰岛新生和胰岛β细胞增殖来增加胰岛质量。抑制β细胞凋亡。
GLP-1 is a potent stimulator of glucose-dependent insulin release. Play important roles on gastric motility and the suppression of plasma glucagon levels. May be involved in the suppression of satiety and stimulation of glucose disposal in peripheral tissues, independent of the actions of insulin. Have growth-promoting activities on intestinal epithelium. May also regulate the hypothalamic pituitary axis (HPA) via effects on LH, TSH, CRH, oxytocin, and vasopressin secretion. Increases islet mass through stimulation of islet neogenesis and pancreatic beta cell proliferaton. Inhibits beta cell apoptosis.
GLP-2刺激小肠的生长并上调小肠的绒毛高度,同时伴有隐窝细胞增殖增加和肠细胞凋亡减少。从胃到结肠的胃肠道是GLP-2作用的主要靶标。在养分稳态,通过增强胃肠功能增强养分吸收以及增加养分处置方面起着关键作用。刺激肠内葡萄糖转运并降低粘膜通透性。
GLP-2 stimulates intestinal growth and up-regulates villus height in the small intestine, concomitant with increased crypt cell proliferation and decreased enterocyte apoptosis. The gastrointestinal tract, from the stomach to the colon is the principal target for GLP-2 action. Plays a key role in nutrient homeostasis, enhancing nutrient assimilation through enhanced gastrointestinal function, as well as increasing nutrient disposal. Stimulates intestinal glucose transport and decreases mucosal permeability.
胃泌酸调节素可显着减少食物摄入。抑制人类的胃排空。抑制胃排空可导致胃扩张增加,这可能通过引起饱胀感而引起饱腹感。
Oxyntomodulin significantly reduces food intake. Inhibits gastric emptying in humans. Suppression of gastric emptying may lead to increased gastric distension, which may contribute to satiety by causing a sensation of fullness.
庆大霉素可以调节胃酸分泌和胃-胃-十二指肠的活性。在生命早期可能对肠道粘膜的生长起重要作用【4】。
Glicentin may modulate gastric acid secretion and the gastro-pyloro-duodenal activity. May play an important role in intestinal mucosal growth in the early period of life【4】.
胰高血糖素(Glucagon)的参考文献:
1. Kieffer TJ and Habener JF (1999). The Glucagon-Like Peptides. Endocrine Reviews, 20 (6), 876-913
2. Jiang G, Zhang BB (2003). Glucagon and regulation of glucose metabolism. Am J Physiol Endocrinol Metab., 284(4), E671-8.
3. Kimball CP, Murlin JR (1923). Aqueous extracts of pancreas. III. Some precipitation reactions of insulin. J. Biol. Chem., 58(1), 337
4. Blache P, Kervran A, Bataille D (1988). Oxyntomodulin and glicentin: brain-gut peptides in the rat. Endocrinology, 123(6), 2782-7.
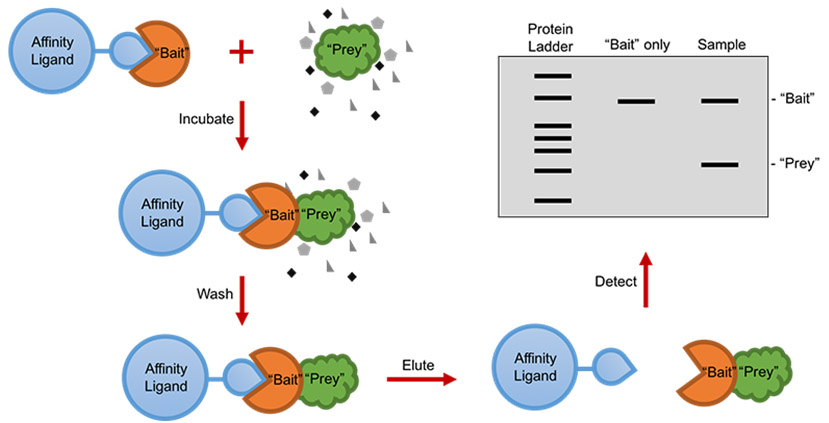
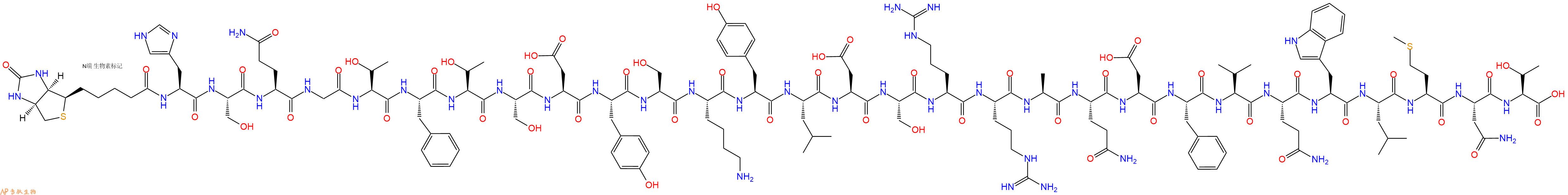
生物素可以与亲和素或者链霉亲和素有力结合,结合强度甚至接近共价键。生物素标记的肽通常用于免疫测定,组织细胞化学和基于荧光的流式细胞术。标记的抗生物素抗体也可以用来结合生物素化多肽。生物素标记常连接在赖氨酸侧链或者N末端。通常在多肽和生物素之间使用6-氨基己酸作为纽带,纽带能够灵活结合底物,并且在有空间位阻的情况下能结合地更好。专肽生物根据需求,向客户提供具有不同位点生物素标记多肽的定制合成。
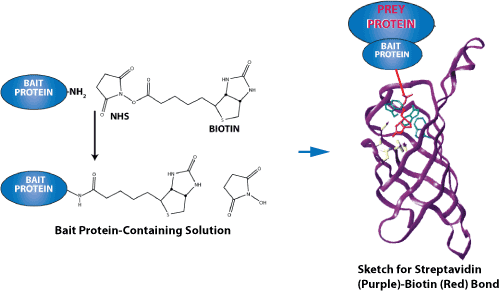
专肽生物合成用于蛋白质-蛋白质相互作用研究的生物素化肽。尽管生物素可以在 N 端或 C 端引入(通过赖氨酸残基),但我们建议使用 N 端修饰,因为它成本低、成功率高、周转时间短且易于操作。因为多肽合成是从 C 端到 N 端合成的,因此,N 端修饰是 SPPS步骤的最后一步,不需要额外的特定缩合步骤。相比之下,C 端修饰需要额外的步骤,并且通常更复杂。当然,原则上生物素可以定位在任何地方。
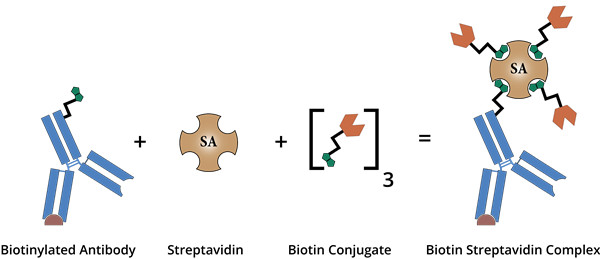
生物素可以通过多种不同的接头或间隔物与肽分离。尽管如此,还是建议包含一个灵活的间隔物,例如 Ahx(一个 6 碳接头),以使生物素标签更加稳定或灵活。
专肽生物在 N 端或 C 端提供生物素化:生物素-N 端、赖氨酸-生物素-肽中间和赖氨酸-生物素-C 端。
专肽生物还可以使用 Ahx 接头或长碳 (LC) 接头提供生物素化:生物素-Ahx-N 末端、Lys-Ahx-生物素-肽中间、Lys-Ahx-生物素-C-末端。
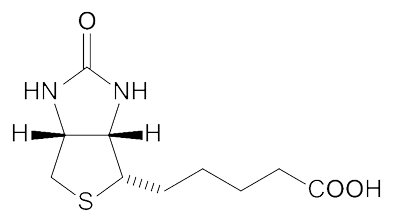
(生物素结构)
示例:
GRGDS在N端和C端标记生物素的结构展示。
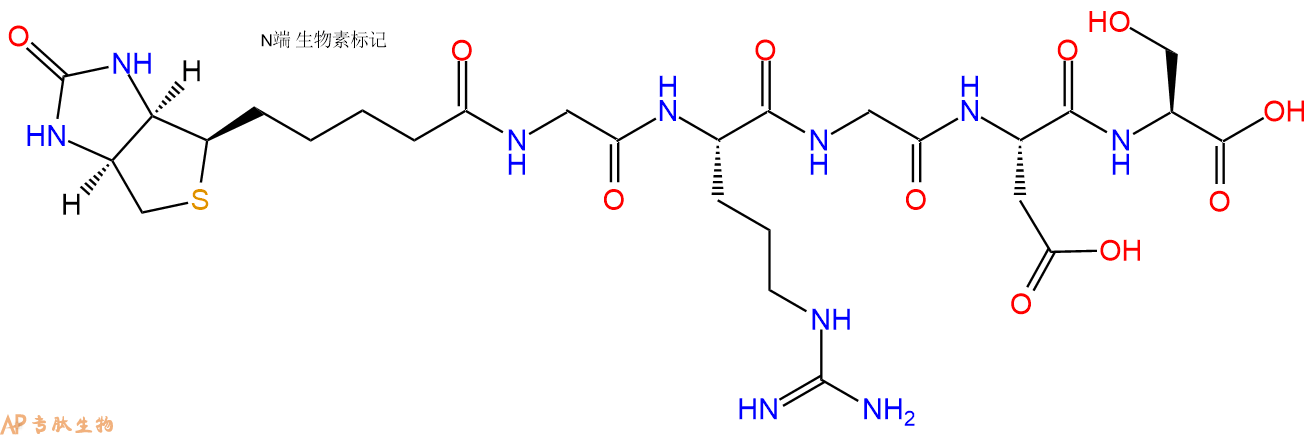
1、GRGDS在N端标记生物素,不增加Ahx 接头
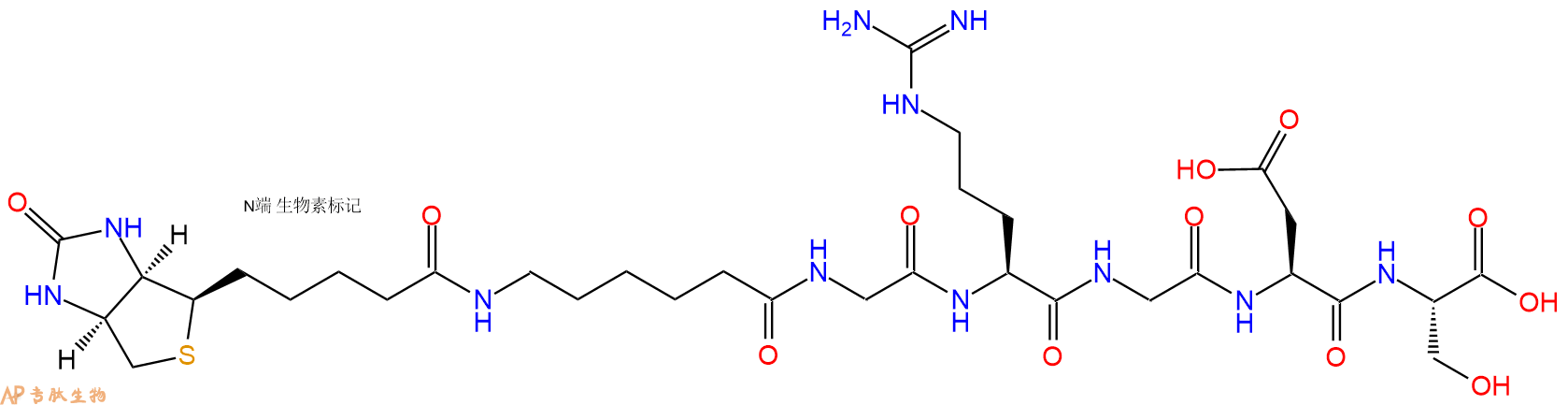
2、GRGDS在N端标记生物素,增加一个Ahx 接头
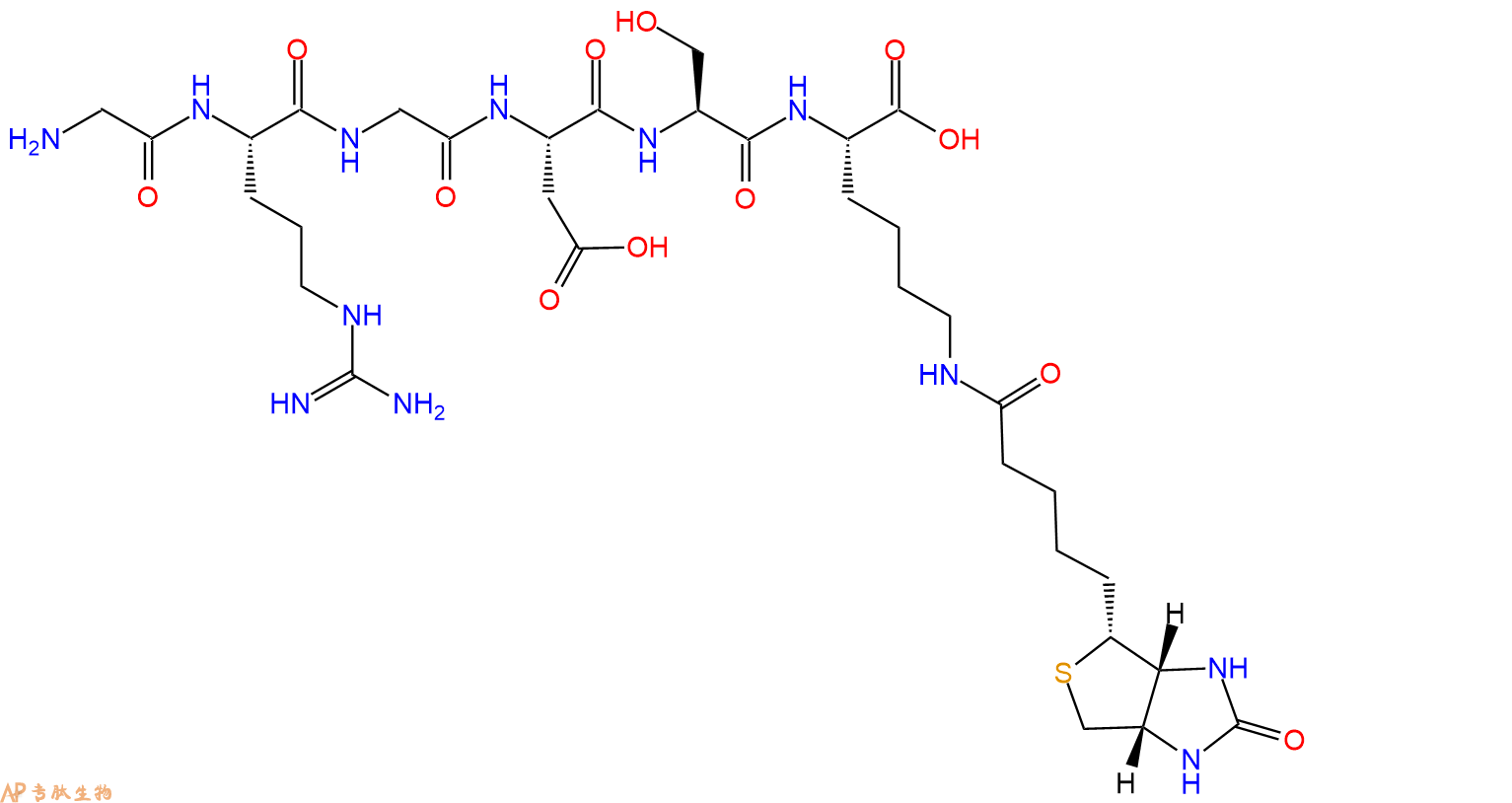
3、GRGDS在C端标记生物素,不增加Ahx 接头
4、GRGDS在C端标记生物素,增加一个Ahx 接头。
A.B.Joshi et al., Int. J. Pharm., 203, 115(2000) : http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=10967434
多肽Biotin-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp-Leu-Met-Asn-Thr-COOH的合成步骤:
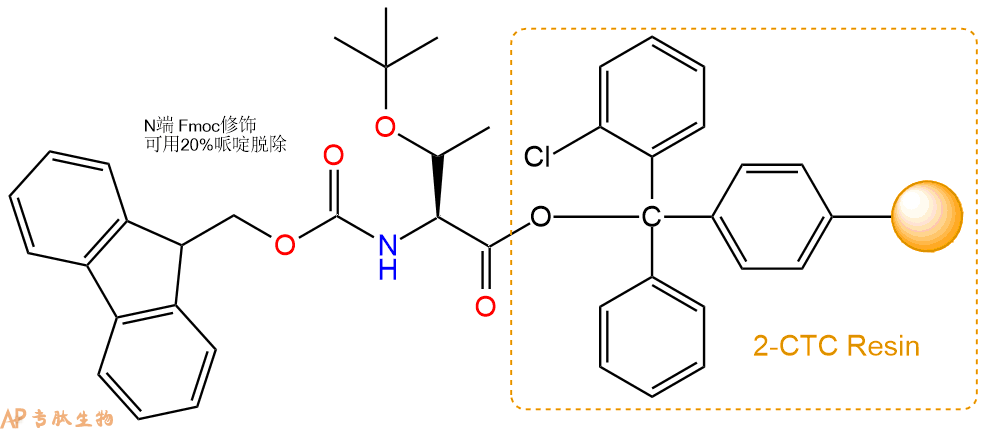
1、合成CTC树脂:称取2.41g CTC Resin(如初始取代度约为0.6mmol/g)和1.74mmol Fmoc-Thr(tBu)-OH于反应器中,加入适量DCM溶解氨基酸(需要注意,此时CTC树脂体积会增大好几倍,避免DCM溶液过少),再加入4.34mmol DIPEA(Mw:129.1,d:0.740g/ml),反应2-3小时后,可不抽滤溶液,直接加入1ml的HPLC级甲醇,封端半小时。依次用DMF洗涤2次,甲醇洗涤1次,DCM洗涤一次,甲醇洗涤一次,DCM洗涤一次,DMF洗涤2次(这里使用甲醇和DCM交替洗涤,是为了更好地去除其他溶质,有利于后续反应)。得到 Fmoc-Thr(tBu)-CTC Resin。结构图如下:
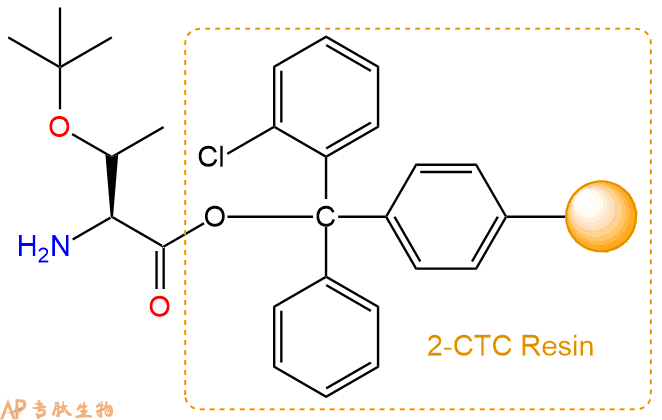
2、脱Fmoc:加3倍树脂体积的20%Pip/DMF溶液,鼓氮气30分钟,然后2倍树脂体积的DMF 洗涤5次。得到 H2N-Thr(tBu)-CTC Resin 。(此步骤脱除Fmoc基团,茚三酮检测为蓝色,Pip为哌啶)。结构图如下:
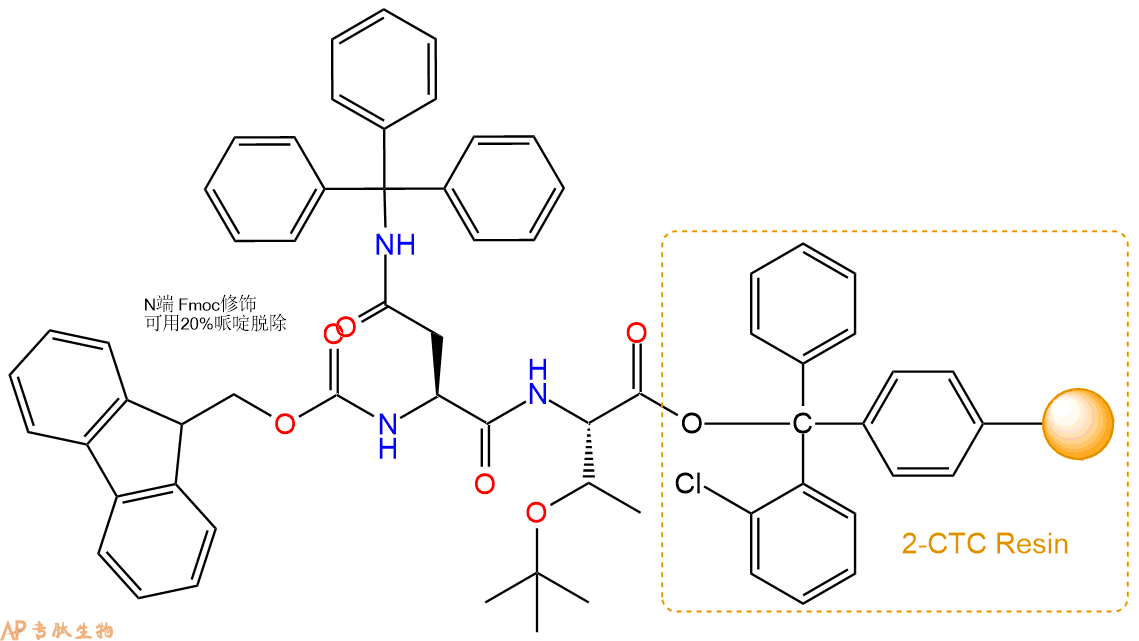
3、缩合:取4.34mmol Fmoc-Asn(Trt)-OH 氨基酸,加入到上述树脂里,加适当DMF溶解氨基酸,再依次加入8.68mmol DIPEA,4.12mmol HBTU。反应30分钟后,取小样洗涤,茚三酮检测为无色。用2倍树脂体积的DMF 洗涤3次树脂。(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。得到Fmoc-Asn(Trt)-Thr(tBu)-CTC Resin。氨基酸:DIPEA:HBTU:树脂=3:6:2.85:1(摩尔比)。结构图如下:
4、依次循环步骤二、步骤三,依次得到
H2N-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Gln(Trt)-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Gln(Trt)-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-Ser(tBu)-Gln(Trt)-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
H2N-Ser(tBu)-Gln(Trt)-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
Fmoc-His(Trt)-Ser(tBu)-Gln(Trt)-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin
以上中间结构,均可在专肽生物多肽计算器-多肽结构计算器中,一键画出。
最后再经过步骤二得到 H2N-His(Trt)-Ser(tBu)-Gln(Trt)-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTC Resin,结构如下:
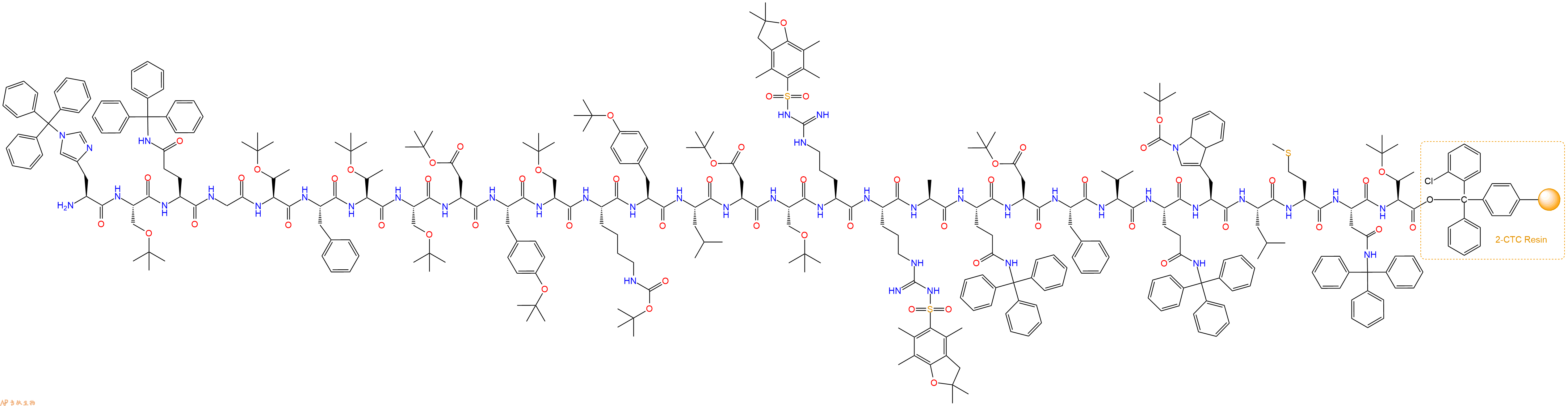
5、生物素反应连接:在上述树脂中,加入适当DMF后,再加入4.34mmol 生物素到树脂中,再加入8.68mmol DIPEA、4.12mmol HBTU,鼓氮气反应30分钟。用2倍树脂体积的DMF 洗涤3次树脂(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。 得到Biotin-His(Trt)-Ser(tBu)-Gln(Trt)-Gly-Thr(tBu)-Phe-Thr(tBu)-Ser(tBu)-Asp(OtBu)-Tyr(tBu)-Ser(tBu)-Lys(Boc)-Tyr(tBu)-Leu-Asp(OtBu)-Ser(tBu)-Arg(Pbf)-Arg(Pbf)-Ala-Gln(Trt)-Asp(OtBu)-Phe-Val-Gln(Trt)-Trp(Boc)-Leu-Met-Asn(Trt)-Thr(tBu)-CTCResin。 结构如下:
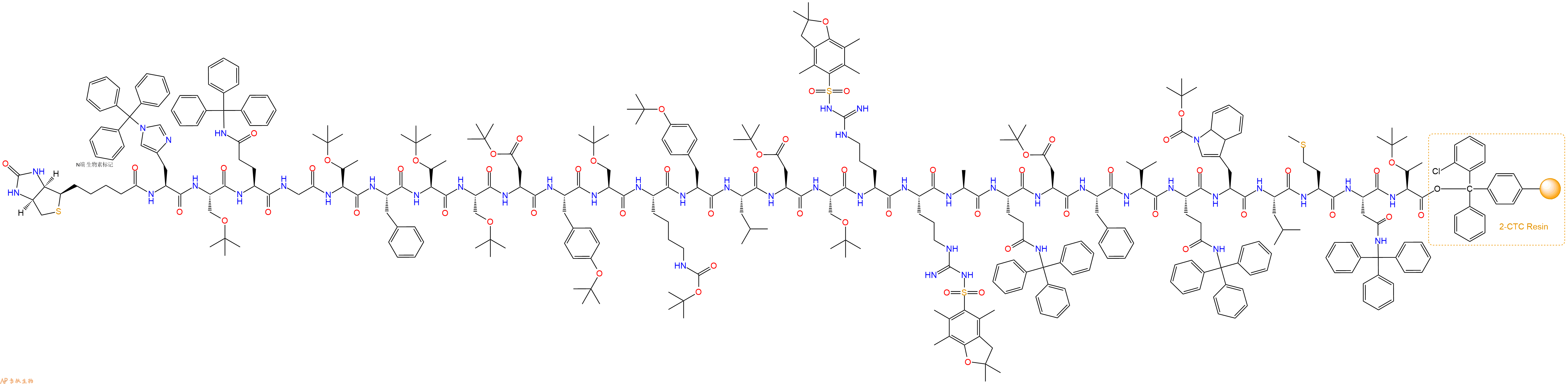
6、切割:6倍树脂体积的切割液(或每1g树脂加8ml左右的切割液),摇床摇晃 2小时,过滤掉树脂,用冰无水乙醚沉淀滤液,并用冰无水乙醚洗涤沉淀物3次,最后将沉淀物放真空干燥釜中,常温干燥24小试,得到粗品Biotin-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp-Leu-Met-Asn-Thr-COOH。结构图见产品结构图。
切割液选择:1)TFA:H2O=95%:5%
2)TFA:H2O:TIS=95%:2.5%:2.5%
3)三氟乙酸:茴香硫醚:1,2-乙二硫醇:苯酚:水=87.5%:5%:2.5%:2.5%:2.5%
(前两种适合没有容易氧化的氨基酸,例如Trp、Cys、Met。第三种适合几乎所有的序列。)
6、纯化冻干:使用液相色谱纯化,收集目标峰液体,进行冻干,获得蓬松的粉末状固体多肽。不过这时要取小样复测下纯度 是否目标纯度。
7、最后总结:
杭州专肽生物技术有限公司(ALLPEPTIDE https://www.allpeptide.com)主营定制多肽合成业务,提供各类长肽,短肽,环肽,提供各类修饰肽,如:荧光标记修饰(CY3、CY5、CY5.5、CY7、FAM、FITC、Rhodamine B、TAMRA等),功能基团修饰肽(叠氮、炔基、DBCO、DOTA、NOTA等),同位素标记肽(N15、C13),订书肽(Stapled Peptide),脂肪酸修饰肽(Pal、Myr、Ste),磷酸化修饰肽(P-Ser、P-Thr、P-Tyr),环肽(酰胺键环肽、一对或者多对二硫键环),生物素标记肽,PEG修饰肽,甲基化修饰肽等。
以上所有内容,为专肽生物原创内容,请勿发布到其他网站上。


















