
Corticotropin-releasing factor human (Human CRF) 能够刺激垂体前叶的促肾上腺皮质激素的合成和分泌。
编号:128610
CAS号:86784-80-7
单字母:H2N-SEEPPISLDLTFHLLREVLEMARAEQLAQQAHSNRKLMEII-NH2
| 编号: | 128610 |
| 中文名称: | 促肾上腺皮质激素释放因子 Corticotropin Releasing Factor CRF human rat |
| 中文同义词: | 促肾上腺皮质激素释放因子醋酸盐 (人) |
| 英文名: | Corticotropin Releasing Factor CRF human rat |
| 英文同义词: | Corticorelin |
| CAS号: | 86784-80-7,醋酸盐 |
| 单字母: | H2N-SEEPPISLDLTFHLLREVLEMARAEQLAQQAHSNRKLMEII-NH2 |
| 三字母: | H2N N端氨基 -Ser丝氨酸 -Glu谷氨酸 -Glu谷氨酸 -Pro脯氨酸 -Pro脯氨酸 -Ile异亮氨酸 -Ser丝氨酸 -Leu亮氨酸 -Asp天冬氨酸 -Leu亮氨酸 -Thr苏氨酸 -Phe苯丙氨酸 -His组氨酸 -Leu亮氨酸 -Leu亮氨酸 -Arg精氨酸 -Glu谷氨酸 -Val缬氨酸 -Leu亮氨酸 -Glu谷氨酸 -Met甲硫氨酸 -Ala丙氨酸 -Arg精氨酸 -Ala丙氨酸 -Glu谷氨酸 -Gln谷氨酰胺 -Leu亮氨酸 -Ala丙氨酸 -Gln谷氨酰胺 -Gln谷氨酰胺 -Ala丙氨酸 -His组氨酸 -Ser丝氨酸 -Asn天冬酰胺 -Arg精氨酸 -Lys赖氨酸 -Leu亮氨酸 -Met甲硫氨酸 -Glu谷氨酸 -Ile异亮氨酸 -Ile异亮氨酸 -NH2C端酰胺化 |
| 氨基酸个数: | 41 |
| 分子式: | C208H344N60O63S2 |
| 平均分子量: | 4757.45 |
| 精确分子量: | 4754.5 |
| 等电点(PI): | 5.58 |
| pH=7.0时的净电荷数: | -1.54 |
| 平均亲水性: | 0.17179487179487 |
| 疏水性值: | -0.26 |
| 外观与性状: | 白色粉末状固体 |
| 消光系数: | - |
| 来源: | 人工化学合成,仅限科学研究使用,不得用于人体。 |
| 纯度: | 95%、98% |
| 盐体系: | 可选TFA、HAc、HCl或其它 |
| 生成周期: | 2-3周 |
| 储存条件: | 负80℃至负20℃ |
| 标签: | ACTH及相关肽 促肾上腺皮质激素释放因子(Corticotropin-Releasing Factor) |
Corticotropin-releasing factor human acetate (Human CRF acetate) 能够刺激垂体前叶的促肾上腺皮质激素的合成和分泌。
Corticotropin-releasing factor human acetate (Human CRF acetate) stimulates the synthesis and secretion of adrenocorticotropin in the anterior pituitary.

ACTH的定义
促肾上腺皮质激素(ACTH、Adrenocorticotropic hormone)或促肾上腺皮质激素是垂体前叶产生的一种激素,它刺激肾上腺皮质。
促肾上腺皮质激素(ACTH),也称为促肾上腺皮质激素,是一种由垂体前叶腺产生和分泌的多肽促性激素。它是下丘脑-垂体-肾上腺轴的重要组成部分,通常是在对生物压力的反应中产生的(连同下丘脑的促肾上腺皮质激素释放激素)。它的主要作用是增加皮质类固醇的产生和释放,以及顾名思义,是肾上腺皮质的皮质醇。
ACTH是由前opiomelanocortin(pre-POMC)合成的。翻译过程中信号肽的去除产生了241个氨基酸的多肽POMC,在被内肽酶进行蛋白水解切割之前,它会经历一系列的翻译后修饰,例如磷酸化和糖基化,以产生具有不同生理活性的各种多肽片段。这些片段包括NPP,促黑素Gamma(γ-MSH),潜在肽,促肾上腺皮质激素(促肾上腺皮质激素或ACTH),促黑素Alpha(促黑素细胞激素或α-MSH),促肾上腺皮质激素样中间肽(CLIP),促肾上腺皮质激素Beta (β-LPH),脂蛋白Gamma(γ-LPH),促黑素β(β-MSH),β-内啡肽和Met-脑啡肽。POMC,
为了调节ACTH的分泌,该轴内分泌的许多物质表现出缓慢/中间和快速的反馈环活性。肾上腺皮质分泌的糖皮质激素起抑制下丘脑CRH分泌的作用,进而降低了ACTH的垂体前叶分泌。糖皮质激素也可能抑制POMC基因转录和肽合成的速率。后者是一个缓慢的反馈循环的例子,它的工作时间从几小时到几天不等,而前者的工作时间则在几分钟到几分钟。
ACTH还与许多生物体的昼夜节律有关。ACTH在人血中的半衰期约为十分钟。
相关肽
六个相关肽包括较小的生物活性片段(激素),这些片段是通过对前opiomelanocortin多蛋白(POMC)进行差分加工而从共同的前体衍生而来的。ACTH,促肾上腺皮质激素样中间叶蛋白[CLIP],β-内啡肽,γ-脂蛋白[yLPH],脑啡肽和α-黑素蛋白[aMSH] 1。
ACTH的发现
1930年代首次研究了ACTH的性质。1933年,由詹姆斯·科利普(James Collip),赫伯特·埃文斯(Herbert Evans)和贝玛尔多·侯赛(Bemardo Houssay)领导的研究小组使用垂体提取物刺激肾上腺皮质。美国生物化学家Choh Hao Li是1943年分离ACTH并在1963年对其进行合成的几位科学家之一。
ACTH的结构特征
ACTH的分子量为4541.3 K Da 2。它是由39个氨基组成的直链肽分子。前24个氨基酸和后7个氨基酸相同,而第25至32个氨基酸略有不同。只有前20个氨基酸才能发挥全部活性,称为活性中心。
ACTH的作用机制
ACTH进入全身循环并与位于肾上腺皮质细胞和皮肤表面的特定高亲和力受体结合。ACTH受体是七种跨膜的G蛋白偶联受体3,在配体结合后会经历构象变化,从而刺激腺苷酸环化酶,从而导致细胞内cAMP的增加和蛋白激酶A的激活。最终导致类固醇生成的刺激。
ACTH的功能
ACTH的主要功能是刺激肾上腺皮质分泌一组类固醇激素,称为糖皮质激素,盐皮质激素和雄激素类固醇。糖皮质激素可控制人体对糖的使用,并在压力时刻帮助调节生物功能。它刺激胆固醇转化为孕烯醇酮,这是所有类固醇激素的前体。它用于治疗类风湿关节炎,溃疡性结肠炎,肝炎和减轻疼痛。它们通过促进必需的蛋白质在特定于记忆的位点4合成所需的蛋白质,在记忆处理中起主要作用。
ACTH的的文献参考
1、Funkelstein L, Toneff T, Mosier C, Hwang SR, Beuschlein F, Lichtenauer UD, Reinheckel T, Peters C, Hook V (2008). Major role of cathepsin L for producing the peptide hormones ACTH, beta-endorphin, and alpha-MSH, illustrated by protease gene knockout and expression. J Biol Chem., 283(51):35652-35659.
2、Lee TH, Lerner AB, Buettner-Janusch V (1961). On the structure of human corticotropin (adrenocorticotropic hormone). J. Biol. Chem., 236:2970-2974.
3、Mountjoy KG, Robbins LS, Mortrud MT, Cone RD (1992). The cloning of a family of genes that encode the melanocortin receptors. Science, 257:248–1251.
4、Flood JF, Jarvik ME, Bennett EL, Orme AE (1976). Effects of ACTH peptide fragments on memory formation. Pharmacol Biochem Behav., 5:41-51.
Definition
Corticotropin-releasing factor (CRF) represents an early chemical signal of the stress response and activates the hypothalamus–pituitary–adrenal axis in response to a stressful stimulus.
Discovery
The existence of CRF was first proposed in 1955 by Guillemin and Rosenberg and Saffran and Schally. Initially isolated from ovine hypothalamus and characterized as a 41 amino acidpeptide in 1981, CRF was subsequently characterizedfrom rat hypothalami, and the identical structure was deduced for human CRF on the basis of the cDNA sequence of the human CRF precursor gene 1,2. Eckart K et al., in 2001 developed strategies permitting synthesis of CRF analogs for CRFBP or CRFR without cross-reaction. They found that one residue of the ARAE motif served as a switch enhancing or preventing ligand binding to CRFBP. This knowledge was applied to the development of peptidic CRF agonists and antagonists 3.
Structural Characteristics
CRF is a 41- amino acid peptide, was originally isolated from the hypothalamus. The stretch of amino acid residues 22–25, Ala-Arg-Ala-Glu, of human/rat CRF, representing the ARAE motif, was found to be responsible for the high affinity of hyrCRF to CRFBP, in contrast to the low affinity of ovine CRF containing the sequence Thr-Lys-Ala-Asp instead. Interestingly, each residue of the ARAE sequence contributed to the high affinity of hyrCRF, as was demonstrated by single residue exchanges. Although sauvagine (Svg), another peptide of the CRF peptide family, binds to CRFBP with only low affinity, the Svg analog containing the ARAE sequence, instead of Glu21-Lys22-Gln23-Glu24, binds with high affinity. However, it has not yet been investigated whether all residues of the ARAE motif are important 4,5. Specific modifications and substitutions of CRF that led to the discovery of antagonists with extended duration of action as compared to that of astressin {cyclo(30−33)[dPhe12,Nle21,Glu30,Lys33,Nle38]hCRF(12-41)}. These additional modifications included elongation of the peptide chain by three residues at the N-terminus, its acetylation, and the [CαMeLeu27] substitution to yield cyclo(30−33)[dPhe12, Nle21,CαMeLeu27,Glu30,Lys33,Nle38]Ac-hCRF(9-41), which was found to be longer acting than astressin which further increases the efficiency (potency, duration of action, and bioavailability) of this family of antagonists 6.
Mode of Action
CRF exhibits its biologic actions through G rotein-dependent receptors. To date, mainly two subtypes of CRF receptors (CRFRs), CRFR1 and CRFR2, have been identified. The central actions of CRF-like peptides are also modulated by the water-soluble CRF binding protein (CRFBP). Consistent with the results of experiments on CRFBP deficient mice, the physiological role of endogenous CRFBP in the central nervous system limit the availability of free ligand for CRFR-mediated actions in brain regions where CRFBP, CRFR, and CRF are colocalized. In addition, central injection of CRF antagonists such as a-helical CRF9–41 (a-hel-CRF9–41), which are bound by CRFBP with high affinity, release endogenous CRF by displacement from CRFBP. The CRF antagonist anti-Svg-30 (aSvg-30) is selective for CRFR2 and useful in in vivo experiments because of its high solubility in cerebrospinal fluid (CSF), the antagonists a-hel-CRF9–41 and Ast are not selective, and bind with different affinities to CRFBP, CRFR1, and CRFR2 7,8.
Functions
Homeostasis, CRF plays a major role in the maintenance or restoration of homeostasis by stimulating the activity of the hypothalamic-pituitary-adrenal (HPA) axis.
Biological function, it also acts within the brain to control immune, reproductive and cardiovascular functions as well as catecholamine release, drug withdrawal, behavior, mood, and anxiety thereby implicating CRF not only as central mediator of stress responses, but also in a variety of stress srelated or -unrelated peripheral functions 9.
On central nervous system, CRF mediates the hypothalamic-pituitary-adrenocortical stress axis. CRF was later found to be widely distributed also outside the hypothalamus throughout the central nervous system 8.
Neurotransmitter, CRF functions as a neurotransmitter or neuromodulator eliciting a wide spectrum of autonomic, electrophysiological, and behavioral effects.
Peripheral sites, in addition to pituitary and central nervous system effects, some effects of CRF in vitro and in vivo have been found at various peripheral sites, where specific binding sites for CRF or messenger RNA (mRNA) for CRF receptors have been localized as well 1.
References
1. Guillemin R, Rosenberg B (1955). Humoral hypothalamic control of anterior pituitary: a study with combined tissue cultures. Endocrinology, 57:599-607.
2. Shibahara S, Morimoto Y, Furutani Y, Notake M, Takahashi H, Shimizu S, Horikawa S, Numa S (1983). Isolation and sequence analysis of the human corticotropin-releasing factor precursor gene. Embo. J., 2(5):775-779.
3. Eckart K, Jahn O, Radulovic J, Tezval H, van Werven L, Spiess J (2001). A single amino acid serves as an affinity switch between the receptor and the binding protein of corticotropin-releasing factor: Implications for the design of agonists and antagonists. PNAS., 98(20):11142-11147.
4. Sutton SW, Behan DP, Lahrichi SL, Kaiser R, Corrigan A, Lowry P, Potter E, Perrin MH, Rivier J, Vale WW (1995). Ligand requirements of the human corticotropin-releasing factor-binding protein. Endocrinology, 136(3):1097-1102.
5. Jahn O, Eckart K, Sydow S, Hofmann BA, Spiess J (2001). Pharmacological characterization of recombinant rat corticotropin releasing factor binding protein using different sauvagine analogs. Peptides, 22(1):47–56.
6. Rivier JE, Kirby DA, Lahrichi SL, Corrigan A, Vale WW, Rivier CL (1999). Constrained corticotropin releasing factor antagonists (astressin analogues) with long duration of action in the rat. J. Med. Chem., 42(16):3175–3182.
7. Spiess J, Dautzenberg FM, Sydow S, Hauger RL, Rühmann A, Blank T, Radulovic J (1998). Molecular Properties of the CRF Receptor. Trends Endocrinol Metab., 9(4):140–145.
8. Brauns O, Liepold T, Radulovic J, Spiess J (2001). Pharmacological and chemical properties of astressin, antisauvagine-30 and alpha-helCRF: significance for behavioral experiments. Neuropharmacology, 41:507–516.
9. Gulyas J, Rivier C, Perrin M, Koerber SC, Sutton S, Corrigan A, Lahrichi SL, Craig AG, Vale W, Rivier J (1995). Potent, structurally constrained agonists and competitive antagonists of corticotropin-releasing factor. PNAS,. 92(23):10575-10579.
Kaneko T et al. Activation of adenylate cyclase-cyclic AMP-protein kinase A signaling by corticotropin-releasing factorwithin the dorsolateral bed nucleus of the stria terminalis is involved in pain-induced aversion. Eur J Neurosci. 2016 Sep 30. : https://pubmed.ncbi.nlm.nih.gov/27690274/
Tenk J et al. Acute central effects of corticotropin-releasing factor (CRF) on energy balance: Effects of age and gender. Peptides. 2016 Nov;85:63-72. : https://pubmed.ncbi.nlm.nih.gov/27637621/
多肽H2N-Ser-Glu-Glu-Pro-Pro-Ile-Ser-Leu-Asp-Leu-Thr-Phe-His-Leu-Leu-Arg-Glu-Val-Leu-Glu-Met-Ala-Arg-Ala-Glu-Gln-Leu-Ala-Gln-Gln-Ala-His-Ser-Asn-Arg-Lys-Leu-Met-Glu-Ile-Ile-NH2的合成步骤:
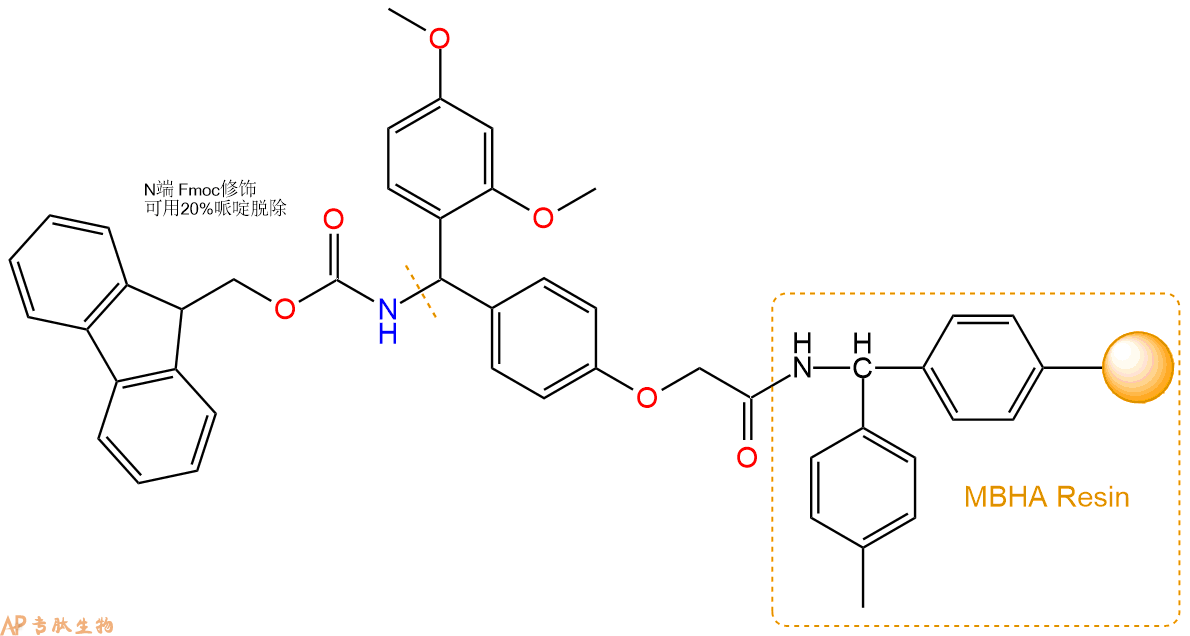
1、合成MBHA树脂:取若干克的MBHA树脂(如初始取代度为0.5mmol/g)和1倍树脂摩尔量的Fmoc-Linker-OH加入到反应器中,加入DMF,搅拌使氨基酸完全溶解。再加入树脂2倍量的DIEPA,搅拌混合均匀。再加入树脂0.95倍量的HBTU,搅拌混合均匀。反应3-4小时后,用DMF洗涤3次。用2倍树脂体积的10%乙酸酐/DMF 进行封端30分钟。然后再用DMF洗涤3次,甲醇洗涤2次,DCM洗涤2次,再用甲醇洗涤2次。真空干燥12小时以上,得到干燥的树脂{Fmoc-Linker-MHBA Resin},测定取代度。这里测得取代度为 0.3mmol/g。结构如下图:
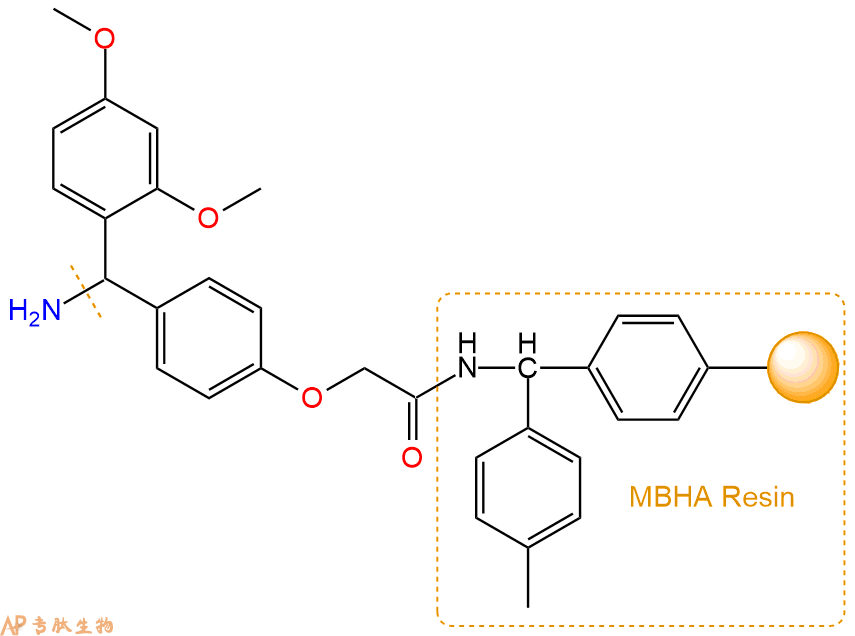
2、脱Fmoc:取1.03g的上述树脂,用DCM或DMF溶胀20分钟。用DMF洗涤2遍。加3倍树脂体积的20%Pip/DMF溶液,鼓氮气30分钟,然后2倍树脂体积的DMF 洗涤5次。得到 H2N-Linker-MBHA Resin 。(此步骤脱除Fmoc基团,茚三酮检测为蓝色,Pip为哌啶)。结构图如下:
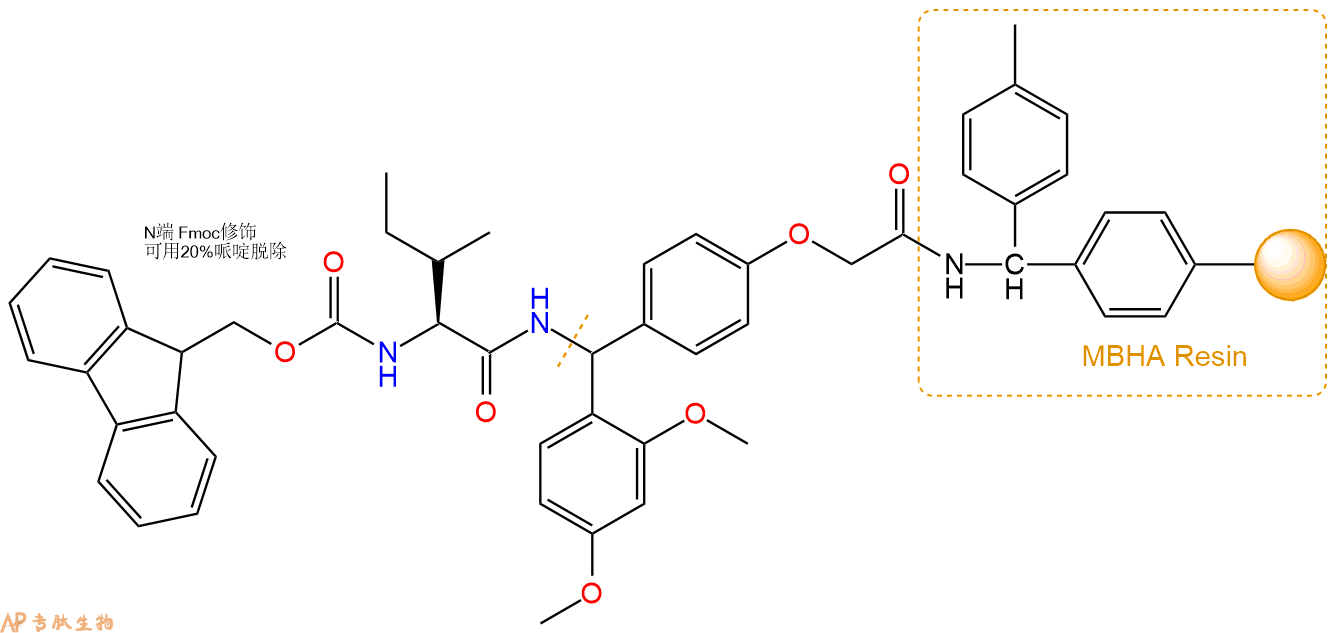
3、缩合:取0.93mmol Fmoc-Ile-OH 氨基酸,加入到上述树脂里,加适当DMF溶解氨基酸,再依次加入1.85mmol DIPEA,0.88mmol HBTU。反应30分钟后,取小样洗涤,茚三酮检测为无色。用2倍树脂体积的DMF 洗涤3次树脂。(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。得到Fmoc-Ile-Linker-MBHA Resin。氨基酸:DIPEA:HBTU:树脂=3:6:2.85:1(摩尔比)。结构图如下:
4、依次循环步骤二、步骤三,依次得到
H2N-Ile-Linker-MBHA Resin
Fmoc-Ile-Ile-Linker-MBHA Resin
H2N-Ile-Ile-Linker-MBHA Resin
Fmoc-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Pro-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Pro-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Glu(OtBu)-Pro-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Glu(OtBu)-Pro-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Glu(OtBu)-Glu(OtBu)-Pro-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
H2N-Glu(OtBu)-Glu(OtBu)-Pro-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
Fmoc-Ser(tBu)-Glu(OtBu)-Glu(OtBu)-Pro-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin
以上中间结构,均可在专肽生物多肽计算器-多肽结构计算器中,一键画出。
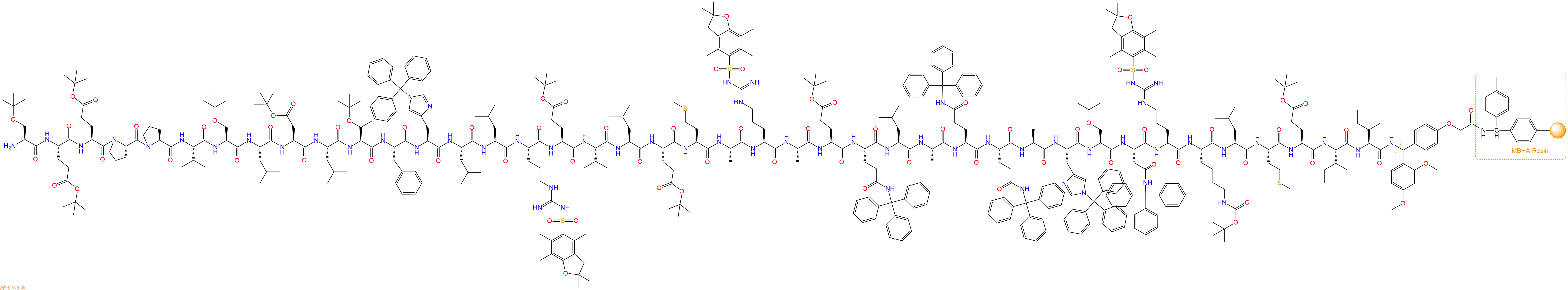
最后再经过步骤二得到 H2N-Ser(tBu)-Glu(OtBu)-Glu(OtBu)-Pro-Pro-Ile-Ser(tBu)-Leu-Asp(OtBu)-Leu-Thr(tBu)-Phe-His(Trt)-Leu-Leu-Arg(Pbf)-Glu(OtBu)-Val-Leu-Glu(OtBu)-Met-Ala-Arg(Pbf)-Ala-Glu(OtBu)-Gln(Trt)-Leu-Ala-Gln(Trt)-Gln(Trt)-Ala-His(Trt)-Ser(tBu)-Asn(Trt)-Arg(Pbf)-Lys(Boc)-Leu-Met-Glu(OtBu)-Ile-Ile-Linker-MBHA Resin,结构如下:
5、切割:6倍树脂体积的切割液(或每1g树脂加8ml左右的切割液),摇床摇晃 2小时,过滤掉树脂,用冰无水乙醚沉淀滤液,并用冰无水乙醚洗涤沉淀物3次,最后将沉淀物放真空干燥釜中,常温干燥24小试,得到粗品H2N-Ser-Glu-Glu-Pro-Pro-Ile-Ser-Leu-Asp-Leu-Thr-Phe-His-Leu-Leu-Arg-Glu-Val-Leu-Glu-Met-Ala-Arg-Ala-Glu-Gln-Leu-Ala-Gln-Gln-Ala-His-Ser-Asn-Arg-Lys-Leu-Met-Glu-Ile-Ile-NH2。结构图见产品结构图。
切割液选择:1)TFA:H2O=95%:5%
2)TFA:H2O:TIS=95%:2.5%:2.5%
3)三氟乙酸:茴香硫醚:1,2-乙二硫醇:苯酚:水=87.5%:5%:2.5%:2.5%:2.5%
(前两种适合没有容易氧化的氨基酸,例如Trp、Cys、Met。第三种适合几乎所有的序列。)
6、纯化冻干:使用液相色谱纯化,收集目标峰液体,进行冻干,获得蓬松的粉末状固体多肽。不过这时要取小样复测下纯度 是否目标纯度。
7、最后总结:
杭州专肽生物技术有限公司(ALLPEPTIDE https://www.allpeptide.com)主营定制多肽合成业务,提供各类长肽,短肽,环肽,提供各类修饰肽,如:荧光标记修饰(CY3、CY5、CY5.5、CY7、FAM、FITC、Rhodamine B、TAMRA等),功能基团修饰肽(叠氮、炔基、DBCO、DOTA、NOTA等),同位素标记肽(N15、C13),订书肽(Stapled Peptide),脂肪酸修饰肽(Pal、Myr、Ste),磷酸化修饰肽(P-Ser、P-Thr、P-Tyr),环肽(酰胺键环肽、一对或者多对二硫键环),生物素标记肽,PEG修饰肽,甲基化修饰肽等。
以上所有内容,为专肽生物原创内容,请勿发布到其他网站上。




















