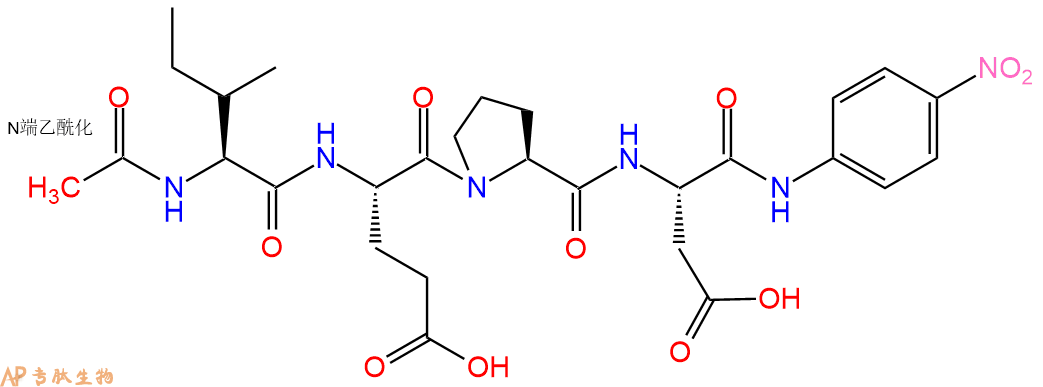
The caspase substrate Ac-IEPD-pNA containing the granzyme B cleavage site was proposed as a potential granzyme B target. It was shown to be the optimal P4-P1 substrate sequence for granzyme B.
编号:161912
CAS号:216757-29-8
单字母:Ac-IEPD-pNA
| 编号: | 161912 |
| 中文名称: | 四肽Ac-Ile-Glu-Pro-Asp-对硝基苯胺 |
| CAS号: | 216757-29-8 |
| 单字母: | Ac-IEPD-pNA |
| 三字母: | Ac N端乙酰化封端 -Ile异亮氨酸 -Glu谷氨酸 -Pro脯氨酸 -Asp天冬氨酸 -pNA对硝基苯胺 |
| 氨基酸个数: | 4 |
| 分子式: | C28H38O11N6 |
| 平均分子量: | 634.63 |
| 精确分子量: | 634.26 |
| 等电点(PI): | - |
| pH=7.0时的净电荷数: | -2 |
| 平均亲水性: | 1.4 |
| 疏水性值: | -1.03 |
| 外观与性状: | 白色粉末状固体 |
| 消光系数: | - |
| 来源: | 人工化学合成,仅限科学研究使用,不得用于人体。 |
| 纯度: | 95%、98% |
| 盐体系: | 可选TFA、HAc、HCl或其它 |
| 储存条件: | 负80℃至负20℃ |
| 标签: | pNA修饰肽 现货多肽 凋亡与坏死 |
The caspase substrate Ac-IEPD-pNA containing the granzyme B cleavage site was proposed as a potential granzyme B target. It was shown to be the optimal P4-P1 substrate sequence for granzyme B.
定义
细胞凋亡或程序性细胞死亡是多细胞生物发育和健康的正常组成部分。细胞响应各种刺激而死亡,在细胞凋亡期间,它们以受控,受控的方式死亡。
发现
1885年, Flemming W描述了程序性细胞死亡的过程。约翰·克尔(John Kerr)在1960年代后期的发现最初被称为“收缩坏死”,但后来改名为“细胞凋亡”,是在他对大鼠急性肝损伤的研究中,他的注意力被好奇的肝细胞死亡形式引起的 1,2。 1972年,Kerr提出了术语“细胞凋亡”的意思是控制细胞缺失的机制,它似乎在调节动物细胞群中与有丝分裂起着互补但相反的作用。它的形态学特征表明它是一种活跃的,固有编程的现象,并且已经表明它可以被多种环境刺激所引发或抑制, 3。
结构特征
Bcl-2家族成员之间的异二聚化是调节程序性细胞死亡的关键事件。通过确定存活蛋白Bcl-xL和Bcl-2相关蛋白Bak的促死亡区域之间的复合物的溶液结构,研究了异二聚体形成的分子基础。突变型Bak肽的结构和结合亲和力表明Bak肽采用两亲性螺旋,通过疏水和静电相互作用与Bcl-xL相互作用。全长Bak的突变会破坏任一类型的相互作用,从而抑制Bak与Bcl-xL 4异源二聚的能力。
通过核磁共振波谱(NMR)确定与Bcl-xL具有生物活性的缺失突变体复合的16-氨基酸肽的结构。由总共2813个NMR约束确定结构,并通过NMR数据很好地定义了结构。当与Bcl-xL复合时,Bak肽形成螺旋。Bak肽的COOH末端部分主要与BH2和BH3区的残基相互作用。黑色素瘤细胞凋亡抑制剂(ML-IAP)是一种有效的抗凋亡蛋白,在许多黑色素瘤细胞系中上调,但在大多数正常成人组织中均未表达。在人类癌症中,IAP蛋白(例如ML-IAP或普遍表达的X染色体连接的IAP(XIAP))的过表达已显示可抑制多种刺激诱导的细胞凋亡。5。
作用方式
一旦收到指示细胞进行凋亡的特定信号,细胞中就会发生许多明显的变化。称为胱天蛋白酶的蛋白质家族通常在凋亡的早期被激活。这些蛋白质分解或切割正常细胞功能所需的关键细胞成分,包括细胞骨架中的结构蛋白和核蛋白(例如DNA修复酶)。半胱天冬酶还可以活化其他降解酶,例如DNase,其开始切割细胞核中的DNA。
凋亡细胞在凋亡过程中表现出独特的形态。通常,在细胞骨架中的lamins和肌动蛋白丝分裂后,细胞开始收缩。染色质在细胞核中的分解通常会导致核浓缩,并且在许多情况下,凋亡细胞的细胞核呈“马蹄形”的外观。细胞继续收缩,将自身包装成可被巨噬细胞去除的形式。有许多机制可以通过诱导细胞凋亡。细胞对这些刺激中任何一种的敏感性可能会因多种因素而异,例如促凋亡和抗凋亡蛋白(例如Bcl-2蛋白或凋亡蛋白的抑制剂)的表达,刺激的严重程度和细胞周期的阶段。Bcl-2蛋白家族在调节多种刺激诱导的凋亡细胞死亡中起着核心作用。该家族中的某些蛋白质,包括Bcl-2和Bcl-xL,可以抑制程序性细胞死亡,而其他蛋白质,例如Bax和Bak,可以促进细胞凋亡 6、7。
功能
对于发育,细胞凋亡与有丝分裂一样是正常发育所必需的。例子:tail变尾时into的吸收被青蛙吸收。
生物体的完整性需要凋亡来破坏对生物体完整性构成威胁的细胞。例子:感染了病毒的细胞8。
免疫系统的细胞,一种细胞介导的免疫反应减弱,必须去除效应细胞以防止它们攻击机体成分。CTLs互相诱导凋亡,甚至自身诱导凋亡9。
具有DNA损伤,破坏其基因组的细胞会导致细胞破坏正常的胚胎发育,导致先天缺陷变成癌。
参考
1. Kerr JF (1965). A histochemical study of hypertrophy and ischaemic injury of rat liver with special reference to changes in lysosomes. Journal of Pathology and Bacteriology, 90(90):419-435.
2. Kerr JF, Wyllie AH, Currie AR (1972). Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer., 26(4):239-257.
3. O'Rourke MG, Ellem KA (2000). John Kerr and apoptosis. Med. J. Aust., 173(11-12): 616-617.
4. Franklin MC, Kadkhodayan S, Ackerly H, Alexandru D, Distefano MD, Elliott LO, Flygare JA, Mausisa G, Okawa DC, Ong D, Vucic D, Deshayes K, Fairbrother WJ (2003). Structure and function analysis of peptide antagonists of melanoma inhibitor of apoptosis (ML-IAP). Biochemistry, 42(27):8223-8231.
5. Sattler M, Liang H, Nettesheim D, Meadows RP, Harlan JE, Eberstadt M, Yoon HS, Shuker SB, Chang BS, Minn AJ, Thompson CB, Fesik SW (1997). Structure of bcl-xl-bak peptide complex: recognition between regulators of apoptosis. Science, 275(5302):983-986.
6. Hanada M, Aimé-Sempé C, Sato T, Reed JC (1995). Structure-function analysis of Bcl-2 protein. Identification of conserved domains important for homodimerization with Bcl-2 and heterodimerization with Bax. J. Biol. Chem., 270(20):11962-11969.
7. Cheng EHY, Levine B, Boise LH, Thompson CB, Hardwic JM (1996). Bax-independent inhibition of apoptosis by Bcl-xL.Nature, 379:554-556.
8. Alimonti JB, Ball TB, Fowke KR (2003). Mechanisms of CD4+ T lymphocyte cell death in human immunodeficiency virus infection and AIDS. J Gen Virology., 84(84): 1649-1661.
9. Werlen G, Hausmann B, Naeher D, Palmer E (2003). Signaling life and death in the thymus: timing is everything. Science. 299(5614):1859-1863.
| DOI | 名称 | |
|---|---|---|
| 10.1074/jbc.273.42.27364 | Definition and redesign of the extended substrate specificity of granzyme B | 下载 |
多肽Ac-Ile-Glu-Pro-Asp-pNA的合成步骤:
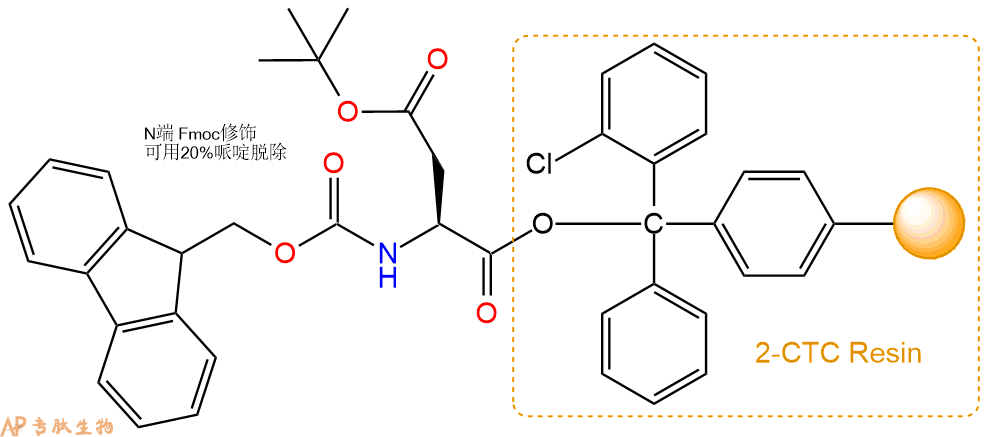
1、合成CTC树脂:称取0.88g CTC Resin(如初始取代度约为0.42mmol/g)和0.44mmol Fmoc-Asp(OtBu)-OH于反应器中,加入适量DCM溶解氨基酸(需要注意,此时CTC树脂体积会增大好几倍,避免DCM溶液过少),再加入1.11mmol DIPEA(Mw:129.1,d:0.740g/ml),反应2-3小时后,可不抽滤溶液,直接加入1ml的HPLC级甲醇,封端半小时。依次用DMF洗涤2次,甲醇洗涤1次,DCM洗涤一次,甲醇洗涤一次,DCM洗涤一次,DMF洗涤2次(这里使用甲醇和DCM交替洗涤,是为了更好地去除其他溶质,有利于后续反应)。得到 Fmoc-Asp(OtBu)-CTC Resin。结构图如下:
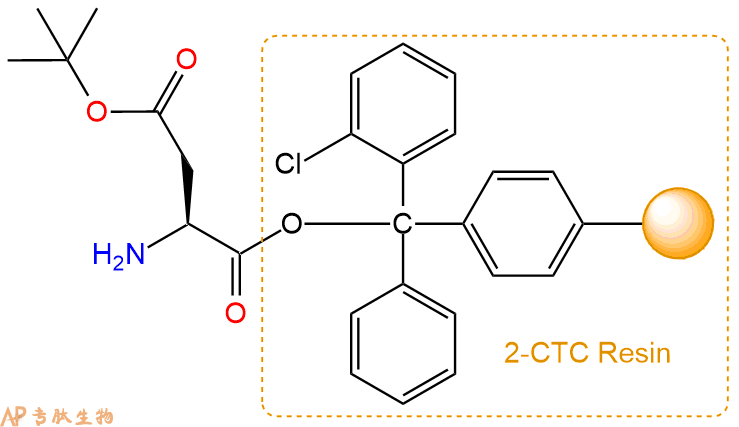
2、脱Fmoc:加3倍树脂体积的20%Pip/DMF溶液,鼓氮气30分钟,然后2倍树脂体积的DMF 洗涤5次。得到 H2N-Asp(OtBu)-CTC Resin 。(此步骤脱除Fmoc基团,茚三酮检测为蓝色,Pip为哌啶)。结构图如下:
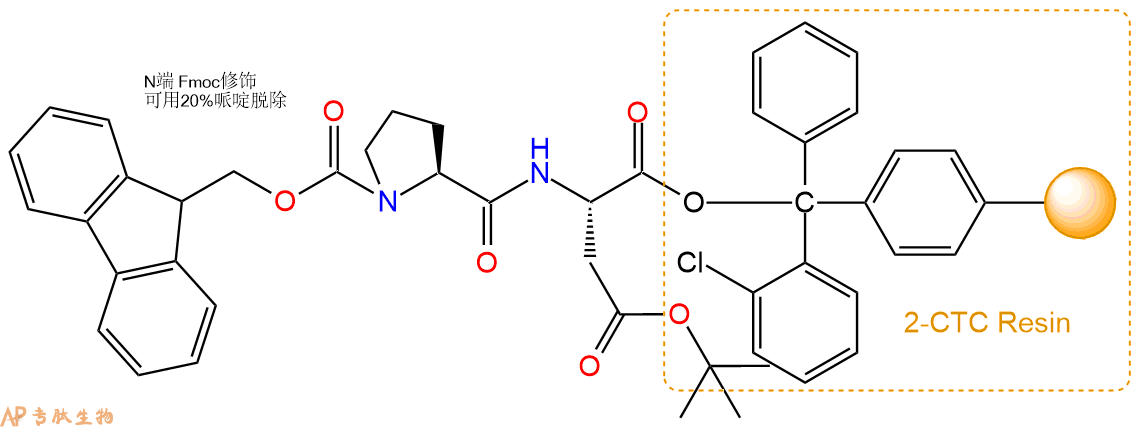
3、缩合:取1.11mmol Fmoc-Pro-OH 氨基酸,加入到上述树脂里,加适当DMF溶解氨基酸,再依次加入2.22mmol DIPEA,1.05mmol HBTU。反应30分钟后,取小样洗涤,茚三酮检测为无色。用2倍树脂体积的DMF 洗涤3次树脂。(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。得到Fmoc-Pro-Asp(OtBu)-CTC Resin。氨基酸:DIPEA:HBTU:树脂=3:6:2.85:1(摩尔比)。结构图如下:
4、依次循环步骤二、步骤三,依次得到
H2N-Pro-Asp(OtBu)-CTC Resin
Fmoc-Glu(OtBu)-Pro-Asp(OtBu)-CTC Resin
H2N-Glu(OtBu)-Pro-Asp(OtBu)-CTC Resin
Fmoc-Ile-Glu(OtBu)-Pro-Asp(OtBu)-CTC Resin
以上中间结构,均可在专肽生物多肽计算器-多肽结构计算器中,一键画出。
最后再经过步骤二得到 H2N-Ile-Glu(OtBu)-Pro-Asp(OtBu)-CTC Resin,结构如下:
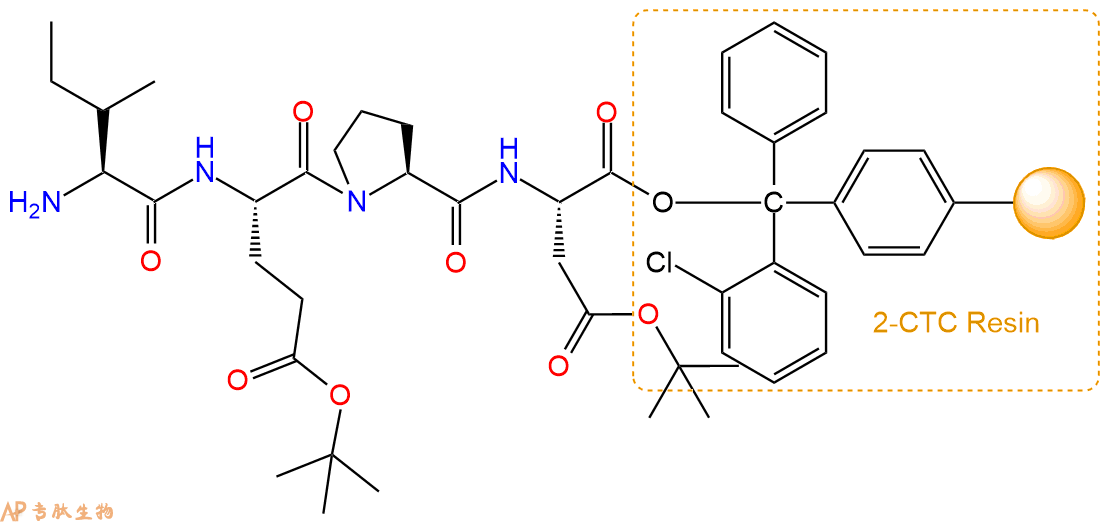
5、乙酸酐反应连接:在上述树脂中,加入适当DMF后,再加入1.11mmol 乙酸酐到树脂中,再加入2.22mmol DIPEA、1.05mmol HBTU,鼓氮气反应30分钟。用2倍树脂体积的DMF 洗涤3次树脂(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。 得到Ac-Ile-Glu(OtBu)-Pro-Asp(OtBu)-CTC Resin。 结构如下:
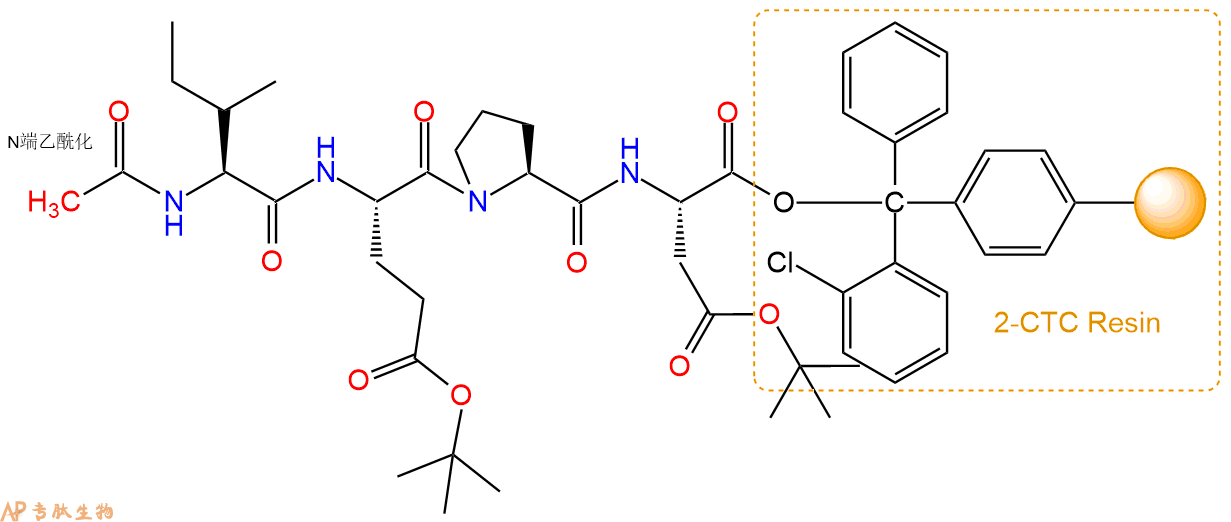
6、全保护切割:配置0.5%TFA/DCM溶液,溶液体积约为树脂体积的3倍。再次用DCM洗涤树脂2遍(去除残留DMF),后将配置好的溶液倒入到反应器中,反应30分钟。抽滤树脂,收集滤液(此时多肽已经从树脂上分离,存在于滤液中)。多肽序列为 Ac-Ile-Glu(OtBu)-Pro-Asp(OtBu)-CTC Resin。 在滤液中添加DIEPA,调PH至7-8。用饱和NaHCO3洗涤滤液,分离出DCM层溶液。可适当旋蒸DCM层溶液,减少有机溶剂。再次加入1或2倍体积的乙酸乙酯,用稀HCl溶液调PH至微酸性,将多肽从DCM层萃取到乙酸乙酯层。用饱和NaCl洗涤2次乙酸乙酯层。用无水硫酸镁吸收乙酸乙酯层的水分。通过减压旋蒸,直接将乙酸乙酯完全旋蒸掉,得到晶体状固体多肽,用于下一步C端反应。或通过减压旋蒸保留适量乙酸乙酯的溶液体积,加入冰乙醚析出 多肽,然后对多肽进行烘干操作即可用于下一步C端反应。Ac-Ile-Glu(OtBu)-Pro-Asp(OtBu)-COOH的结构图如下。
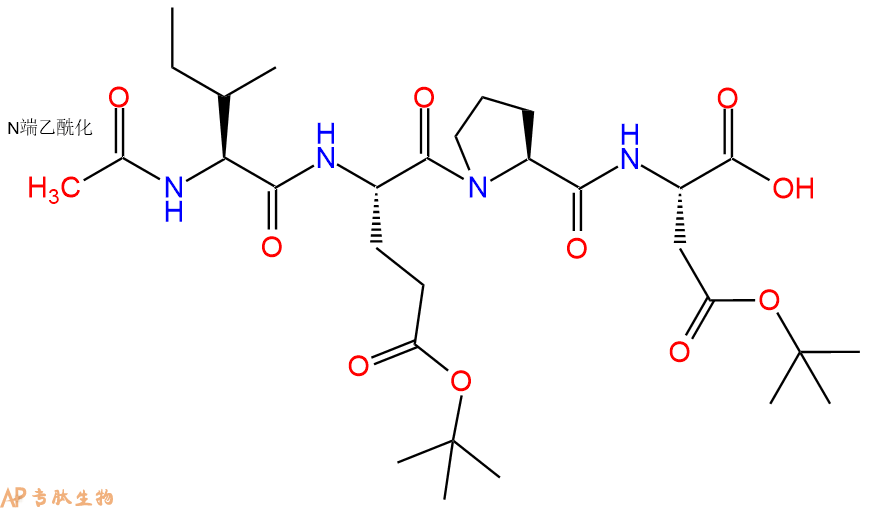
7、4-硝基苯胺反应连接:在上述树脂中,加入适当DMF后,再加入1.11mmol 4-硝基苯胺到树脂中,再加入2.22mmol DIPEA、1.05mmol HBTU,鼓氮气反应30分钟。用2倍树脂体积的DMF 洗涤3次树脂(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。 得到 Ac-Ile-Glu(OtBu)-Pro-Asp(OtBu)-pNA。 结构如下:
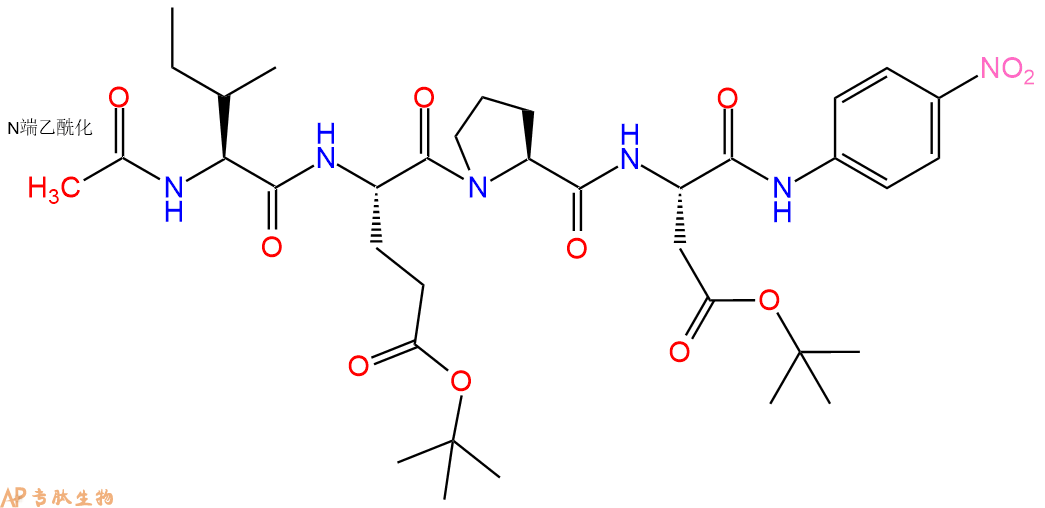
8、切割:6倍树脂体积的切割液(或每1g树脂加8ml左右的切割液),摇床摇晃 2小时,过滤掉树脂,用冰无水乙醚沉淀滤液,并用冰无水乙醚洗涤沉淀物3次,最后将沉淀物放真空干燥釜中,常温干燥24小试,得到粗品Ac-Ile-Glu-Pro-Asp-pNA。结构图见产品结构图。
切割液选择:1)TFA:H2O=95%:5%
2)TFA:H2O:TIS=95%:2.5%:2.5%
3)三氟乙酸:茴香硫醚:1,2-乙二硫醇:苯酚:水=87.5%:5%:2.5%:2.5%:2.5%
(前两种适合没有容易氧化的氨基酸,例如Trp、Cys、Met。第三种适合几乎所有的序列。)
9、纯化冻干:使用液相色谱纯化,收集目标峰液体,进行冻干,获得蓬松的粉末状固体多肽。不过这时要取小样复测下纯度 是否目标纯度。
10、最后总结:
杭州专肽生物技术有限公司(ALLPEPTIDE https://www.allpeptide.com)主营定制多肽合成业务,提供各类长肽,短肽,环肽,提供各类修饰肽,如:荧光标记修饰(CY3、CY5、CY5.5、CY7、FAM、FITC、Rhodamine B、TAMRA等),功能基团修饰肽(叠氮、炔基、DBCO、DOTA、NOTA等),同位素标记肽(N15、C13),订书肽(Stapled Peptide),脂肪酸修饰肽(Pal、Myr、Ste),磷酸化修饰肽(P-Ser、P-Thr、P-Tyr),环肽(酰胺键环肽、一对或者多对二硫键环),生物素标记肽,PEG修饰肽,甲基化修饰肽
以上所有内容,为专肽生物原创内容,请勿发布到其他网站上。




















