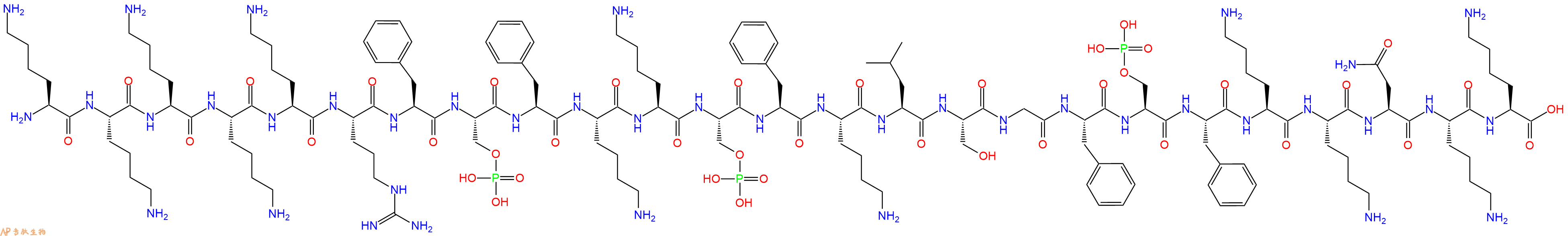
一种由豆蔻酰化富丙氨酸 C 激酶底物蛋白 (MARCKS) 基本效应域片段组成的磷酸化多肽,MARCKS Peptide (151-175) 的磷酸化逆转了它对磷脂酶 C (PLC) 催化磷脂酰肌醇 -4,5- 二磷酸 (PIP2) 水解的抑制作用。
编号:153313
CAS号:
单字母:H2N-KKKKKRF-pSer-FKK-pSer-FKLSGF-pSer-FKKNKK-OH
| 编号: | 153313 |
| 中文名称: | MARCKSSubstrate(151-175) |
| 英文名: | MARCKSSubstrate(151-175) |
| 单字母: | H2N-KKKKKRF-pSer-FKK-pSer-FKLSGF-pSer-FKKNKK-OH |
| 三字母: | H2N N端氨基 -Lys赖氨酸 -Lys赖氨酸 -Lys赖氨酸 -Lys赖氨酸 -Lys赖氨酸 -Arg精氨酸 -Phe苯丙氨酸 -Ser(PO3H2)磷酸化丝氨酸 -Phe苯丙氨酸 -Lys赖氨酸 -Lys赖氨酸 -Ser(PO3H2)磷酸化丝氨酸 -Phe苯丙氨酸 -Lys赖氨酸 -Leu亮氨酸 -Ser丝氨酸 -Gly甘氨酸 -Phe苯丙氨酸 -Ser(PO3H2)磷酸化丝氨酸 -Phe苯丙氨酸 -Lys赖氨酸 -Lys赖氨酸 -Asn天冬酰胺 -Lys赖氨酸 -Lys赖氨酸 -OHC端羧基 |
| 氨基酸个数: | 25 |
| 分子式: | C147H246N41O40P3 |
| 平均分子量: | 3320.7 |
| 精确分子量: | 3318.77 |
| 等电点(PI): | - |
| pH=7.0时的净电荷数: | 13.97 |
| 平均亲水性: | 1.0875 |
| 疏水性值: | -1.62 |
| 外观与性状: | 白色粉末状固体 |
| 消光系数: | - |
| 来源: | 人工化学合成,仅限科学研究使用,不得用于人体。 |
| 纯度: | 95%、98% |
| 盐体系: | 可选TFA、HAc、HCl或其它 |
| 生成周期: | 2-3周 |
| 储存条件: | 负80℃至负20℃ |
| 标签: | 磷酸化修饰肽 酶底物肽(Substrate Peptide) |
MARCKS Peptide(151-175), Phosphorylated 是一种由豆蔻酰化富丙氨酸 C 激酶底物蛋白 (MARCKS) 基本效应域片段组成的磷酸化多肽,MARCKS Peptide (151-175) 的磷酸化逆转了它对磷脂酶 C (PLC) 催化磷脂酰肌醇 -4,5- 二磷酸 (PIP2) 水解的抑制作用。
MARCKS Peptide(151-175), Phosphorylated is a phosphorylated peptide corresponding to the basic effector domain of myristoylated alanine-rich protein kinase C substrate protein (MARCKS). Phosphorylation of MARCKS Peptide (151-175) reverses its inhibition of phospholipase C (PLC)-catalyzed hydrolysis of phosphatidylinositol 4,5-bisphosphate (PIP2)[1].
磷酸肽
在生命过程中发挥重要作用,磷酸化的位置在多肽上的Tyr、Ser,Thr,。目前磷酸肽合成一般都采用磷酸化氨基酸,目前使用的都是单苄基磷酸化氨基酸。磷酸化氨基酸的连接一般采用HBTU/HOBt/DIEA方法,但是目前采用该方法合成磷酸化多肽也有缺点,特别是在合成多磷酸化多肽或氨基酸较长的多肽的时候,连接效率低,最后产品纯度很低,对于这种磷酸化多肽,我们考虑采用后磷酸化方法,其合成过程就是在多肽合成结束后,选择性脱去要标记的氨基酸的侧链保护基,对于Tyr,Thr可以直接使用侧链不保护的氨基酸进行反应,而Ser可以采用Fmoc-Ser(trt),在1% TFA/DCM条件下可以定量的脱除。
后磷酸化,采用双苄基亚磷酰胺,四氮唑生成亚磷酰胺四唑活性中间体,连接到羟基上,随后在过氧酸下氧化生成磷酰基,完成反应。
目前,多肽的磷酸化修饰方法主要有两种:
(1)将适当保护的磷酸化氨基酸直接引入到多肽序列中;
(2)多肽序列在树脂上合成完后,再对其中的Ser、Tyr或Thr的侧链羟基进行磷酸化。
1)将适当保护的磷酸化氨基酸直接引入到多肽序列中:
即事先将需要磷酸化的氨基酸(Thr,Ser或Tyr)磷酸化并适当保护,然后按照正常SPPS 合成流程将磷酸化单体缩合到多肽指定位点。这种方法操作简便,已经成为多肽单位点 磷酸化修饰的主要方法。
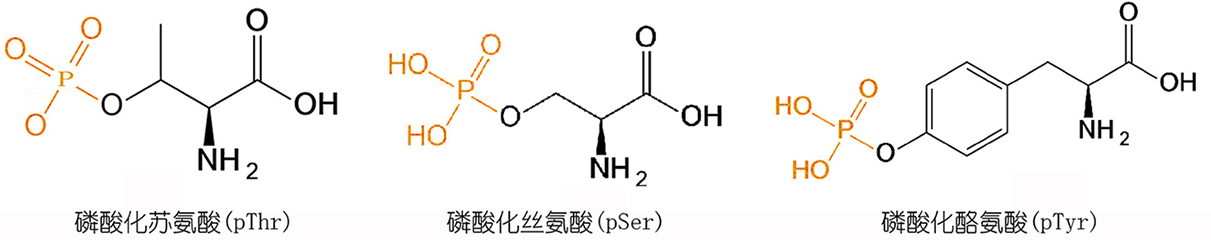
2)多肽序列在树脂上合成完后,再对其中的Ser、Tyr或Thr的侧链羟基进行磷酸化:
采用将磷酸化单体缩合到多肽中的方法进行磷酸化修饰时,磷酸化的氨基酸由于侧链修 饰的较大基团产生的位阻而导致难以与肽链缩合,并且之后的氨基酸引入都会比较困难, 尤其在含有多个磷酸化位点修饰时,合成将变得异常困难,并且最终产物成分复杂,难 以分离,产率极低。
因此,当肽链中多个位点进行磷酸化时,可以考虑采用将多肽序列 在树脂上合成完后,再对其中的Ser、Tyr或Thr的侧链羟基进行磷酸化:其合成过程主要 就是在多肽合成结束之后,选择性的脱去要标记氨基酸的侧链保护基,对于Tyr,Thr可 以直接使用侧链不保护的氨基酸进行反应。
侧链保护基在1%TFA/DCM条件下可以定量的脱 除。采用这种方法时,可以采用双苄基亚磷酰胺,四氮唑生成亚磷酰胺四唑活性中间体, 连接到羟基上,然后在过氧酸条件下氧化生成磷酰基,完成反应。
Caspase酶对应的底物,Caspases(半胱氨酸天冬氨酸蛋白酶,半胱氨酸依赖性天冬氨酸定向蛋白酶)是一类蛋白酶家族,其功能与凋亡(程序性细胞死亡),坏死和发烧(炎症)的过程密切相关。
什么是胱天蛋白酶?
胱天蛋白酶(Caspases)是含半胱氨酸的天冬氨酸蛋白水解酶,它们是为细胞凋亡的主要介质。多种受体,例如TNF-α 受体,FasL受体,TLR和死亡受体,以及Bcl-2和凋亡抑制剂(IAP)蛋白家族参与并调节该caspase依赖性凋亡途径。一旦Caspase受到上游信号(外部或内在)刺激被激活,即会参与执行下游蛋白底物的水解作用,并触发一系列事件,导致细胞分解,死亡,吞噬作用和细胞碎片的清除。
人Caspases酶
人的Caspases家族基于序列相似性和生物学功能等共性主要可分为三大类:第一类由具有长胱天蛋白酶募集结构域的“炎症”胱天蛋白酶组成,他们对P4位上的较大的芳香族或疏水性残基具有亲和力。第二类由具有短的前体结构域的“细胞凋亡效应”胱天蛋白酶组成,而第三类由具有长的前提结构域的Pap位置具有亮氨酸或缬氨酸底物亲和力的“凋亡引发剂”胱天蛋白酶组成(表1)。
表1. 人胱天蛋白酶的功能分类:
| 细胞死亡途径 | 半胱天冬酶类型 | 酵素 | 物种 |
| 细胞凋亡 | 启动器 | Caspases 2 | 人与鼠 |
| 细胞凋亡 | 启动器 | Caspases 8 | 人与鼠 |
| 细胞凋亡 | 启动器 | Caspases 9 | 人与鼠 |
| 细胞凋亡 | 启动器 | Caspases 10 | 人的 |
| 细胞凋亡 | 效应器 | Caspases 3 | 人与鼠 |
| 细胞凋亡 | 效应器 | Caspases 6 | 人与鼠 |
| 细胞凋亡 | 效应器 | Caspases 6 | 人与鼠 |
| 细胞焦亡 | 炎性的 | Caspases 1 | 人与鼠 |
| 细胞焦亡 | 炎性的 | Caspases 4 | 人的 |
| 细胞焦亡 | 炎性的 | Caspases 5 | 人的 |
启动器Caspase和效应器Caspase酶
根据其在凋亡胱天蛋白酶途径中的作用,胱天蛋白酶可分为两类:启动器和效应器Caspase酶。启动器和效应器Caspas酶都具有由小亚基和大亚基组成的催化位点,Caspase酶的识别位
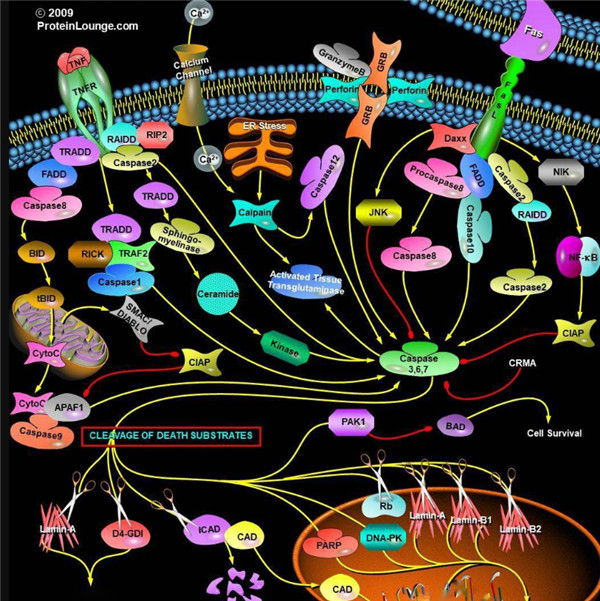
凋亡启动器Caspase酶,例如caspase-2,-8,-9和-10可以启动caspase激活级联反应。Caspase-8对于形成死亡诱导信号复合物(DISC)是必不可少的,并且在激活后,Caspase-8激活下游效应子Caspase(例如Caspase 3)并介导线粒体中细胞色素c的释放。Caspase-8已被证明对IETD肽序列具有相对较高的底物选择性。凋亡效应胱天蛋白酶例如Caspase-3,-6和-7虽然不负责启动级联途径,但是当被激活时,它们在级联的中间和后续步骤中起着不可或缺的作用。Caspase-3(CPP32 / apopain)是关键效应器,因为它放大了来自启动器Caspase的信号,使用对Caspase-3有选择性的DEVD肽序列对活化的Caspase-3进行检测,可以检测Caspase-3的活性。
Caspase酶底物和抑制剂
Caspase底物和抑制剂由两个关键成分组成:Caspase识别序列和信号产生或蛋白酶抑制基序。不同Caspase识别序列不同,一般由三个或四个氨基酸组成(表2)。Caspase酶识别序列的N端通常有乙酰基(Ac)或碳苯甲氧基(Z)基团修饰,以增强膜的通透性。对应的Caspase识别特定的肽序列为其酶促反应切割位点,释放产生信号或抑制信号的基序。Caspase的显色和荧光底物均以相似的方式起作用,其中底物的信号或颜色强度与蛋白水解活性成正比。
表2. Caspase的底物及其序列
| 多肽 | 氨基酸序列 | 对应的Caspase的种类 |
| IETD | Ile-Glu-Thr-Asp | Caspase 8,颗粒酶B |
| DEVD | Asp-Glu-Val-Asp | Caspase 3、6、7、8或10 |
| LEHD | Leu-Glu-His-Asp | Caspase 9 |
| VAD | Val-Ala-Asp | Caspase 1、2、3、6、8、9或10 |
Caspase酶的显色底物
Caspase的显色底物是有Caspase识别序列及生色基团组成,常见的生色团有pNA(对硝基苯胺或4-硝基苯胺),可使用酶标仪或分光光度计在405 nm处进行光密度检测。
表3. Caspase的显色底物
| 底物 | Caspase | 吸收(nm) | 颜色 |
| Ac-DEVD-pNA * CAS 189950-66-1 * | 半胱天冬酶3 | 405 nm | 黄色 |
| Z-DEVD-pNA | 半胱天冬酶3 | 405 nm | 黄色 |
| Z-IETD-pNA * CAS 219138-21-3 * | 半胱天冬酶8,颗粒酶B | 405 nm | 黄色 |
Caspase的荧光底物
Caspase的荧光底物的结构包含与半胱天冬酶识别相关的荧光团,例如7-氨基-4-甲基香豆素(AMC),7-氨基-4-三氟甲基香豆素(AFC), Rhodamine 110(R110)或ProRed™620。R110的Caspase底物比基于香豆素的Caspase底物(例如AMC和AFC)更敏感,但由于两步裂解过程,其动态范围更窄。 建议将R110标记的Caspase底物用于终点法测定,而将AMC和AFC标记的 Caspase底物用于动力学测定。
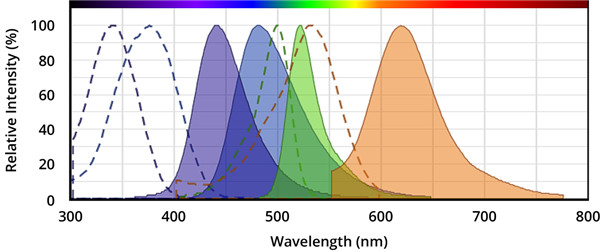
图.从左到右,分别是AMC(7-氨基-4-甲基香豆素),AFC(7-氨基-4-三氟甲基香豆素),Rhodamine 110(R110)和ProRed™620的激发和发射光谱。
表4.荧光半胱天冬酶底物。
| 底物名称 | 对应的Caspase | Ex(nm) | Em(nm) | ε¹ | Φ² |
| Ac-DEVD-AFC * CAS 201608-14-2 * | 半胱天冬酶3、7 | 376 | 482 | 17000 | 0.53 |
| Ac-DEVD-AMC * CAS 169332-61-0 * | 半胱天冬酶3、7 | 341 | 441 | 19000 | N / D |
| Z-DEVD-AFC | 半胱天冬酶3、7 | 376 | 482 | 17000 | 0.53 |
| Z-DEVD-AMC * CAS 1135416-11-3 * | 半胱天冬酶3、7 | 341 | 441 | 19000 | N / D |
| Z-DEVD-ProRed™620 | 半胱天冬酶3、7 | 532 | 619 | N / D | N / D |
| (Z-DEVD)2 -R110 * CAS 223538-61-2 * | 半胱天冬酶3、7 | 500 | 522 | 80000 | N / D |
| Z-DEVD-ProRed™620 | 半胱天冬酶3、7 | 532 | 619 | N / D | N / D |
| Ac-IETD-AFC * CAS 211990-57-7 * | 半胱天冬酶8,颗粒酶B | 376 | 482 | 17000 | 0.53 |
| Z-IETD-AFC * CAS 219138-02-0 * | 半胱天冬酶8,颗粒酶B | 376 | 482 | 17000 | 0.53 |
注意:
1.ε=在其最大吸收波长处的摩尔消光系数(单位= cm -1M -1)。
2.Φ=水性缓冲液(pH 7.2)中的荧光量子产率。
Caspase抑制剂
Caspase抑制剂能与Caspase的活性位点结合并形成可逆或不可逆的连接,通常,Caspase抑制剂的结构由Caspase识别序列,诸如醛(-CHO)或氟甲基酮(-FMK)的官能团组成。具有醛官能团的胱天蛋白酶抑制剂是可逆的,而具有FMK的抑制剂是不可逆的。半胱天冬酶底物和抑制剂都具有较小的细胞毒性作用,因此,它们是研究半胱天冬酶活性的有用工具。
表5. 可逆和不可逆的Caspase酶抑制剂
| 抑制剂 | Caspase的种类 | 是否可逆 | Ex(nm) | Em(nm) |
| Ac-DEVD-CHO * CAS 169332-60-9 * | 半胱天冬酶3、7 | 可逆的 | -- | -- |
| Ac-IETD-CHO * CAS 191338-86-0 * | 半胱天冬酶8 | 可逆的 | -- | -- |
| mFluor™450-VAD-FMK | 半胱天冬酶1,2,3,6,8,9,10 | 不可逆的 | 406 | 445 |
| mFluor™510-VAD-FMK | 半胱天冬酶1,2,3,6,8,9,10 | 不可逆的 | 412 | 505 |
| FITC-C6-DEVD-FMK | 半胱天冬酶3、7 | 不可逆的 | 491 | 516 |
| FITC-C6-DEVD-FMK | 半胱天冬酶3、7 | 不可逆的 | 491 | 516 |
| FITC-C6-LEHD-FMK | 半胱天冬酶9 | 不可逆的 | 491 | 516 |
| FITC-C6-LEHD-FMK | 半胱天冬酶9 | 不可逆的 | 491 | 516 |
| FAM-VAD-FMK | 半胱天冬酶1,2,3,6,8,9,10 | 不可逆的 | 493 | 517 |
| SRB-VAD-FMK [磺胺丁胺B-VAD-FMK] | 半胱天冬酶1,2,3,6,8,9,10 | 不可逆的 | 559 | 577 |
| DOI | 名称 | |
|---|---|---|
| 10.1074/jbc.271.42.26187 | Myristoylated alanine-rich C kinase substrate (MARCKS) produces reversible inhibition of phospholipase C by sequestering phosphatidylinositol 4,5-bisphosphate in lateral domains | 下载 |




















