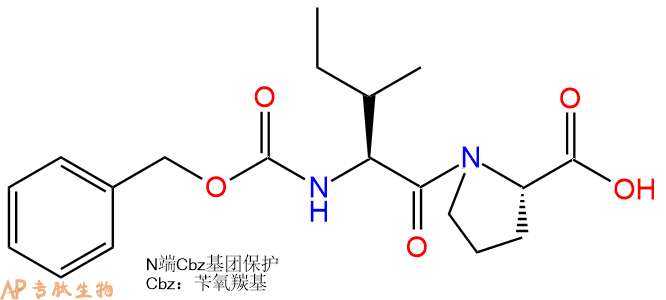
| 编号: | 200440 |
| 中文名称: | 二肽Z-Ile-Pro |
| CAS号: | 13211-37-5 |
| 单字母: | Z-IP-OH |
| 三字母: | Cbz N端Cbz保护 -Ile异亮氨酸 -Pro脯氨酸 -OHC端羧基 |
| 氨基酸个数: | 2 |
| 分子式: | C19H26O5N2 |
| 平均分子量: | 362.42 |
| 精确分子量: | 362.18 |
| 等电点(PI): | - |
| pH=7.0时的净电荷数: | - |
| 平均亲水性: | -1.8 |
| 疏水性值: | 1.45 |
| 消光系数: | - |
| 来源: | 人工化学合成,仅限科学研究使用,不得用于人体。 |
| 储存条件: | 负80℃至负20℃ |
| 标签: | 抑制剂相关肽(Inhibitor Peptide) 淀粉样肽(Amyloid Peptides) CBZ修饰肽 二肽(Dipeptide) |
Z-IL-CHO, a cell-permeable, reversible inhibitor, reduces amyloid β (Aβ)-production by blocking γ-secretase activity in vitro (Aβ₄₀ IC₅₀ = 7.9 µM; Aβ₄₂ IC₅₀ = 7.6 µM). Besides blocking the formation of Aβ the dipeptide aldehyde Z-IL-CHO also blocks the production of the γ-CTF (γ-secretase-generated carboxyl-terminal fragment), but does not affect the formation of amyloid-β-precursor protein (APP).
定义
酶是用于生化反应的非常有效的催化剂。它们通过提供较低活化能的替代反应途径来加快反应速度。酶作用于底物并产生产物。一些物质降低或什至停止酶的催化活性被称为抑制剂。
发现
1965年,Umezawa H分析了微生物产生的酶抑制剂,并分离出了抑制亮肽素和抗痛药的胰蛋白酶和木瓜蛋白酶,乳糜蛋白酶抑制的胰凝乳蛋白酶,胃蛋白酶抑制素抑制胃蛋白酶,泛磷酰胺抑制唾液酸酶,乌藤酮抑制酪氨酸羟化酶,多巴汀抑制多巴胺3-羟硫基嘧啶和多巴胺3-羟色胺酶酪氨酸羟化酶和多巴胺J3-羟化酶。最近,一种替代方法已应用于预测新的抑制剂:合理的药物设计使用酶活性位点的三维结构来预测哪些分子可能是抑制剂1。已经开发了用于识别酶抑制剂的基于计算机的方法,例如分子力学和分子对接。
结构特征
已经确定了许多抑制剂的晶体结构。已经确定了三种与凝血酶复合的高效且选择性的低分子量刚性肽醛醛抑制剂的晶体结构。这三种抑制剂全部在P3位置具有一个新的内酰胺部分,而对胰蛋白酶选择性最高的两种抑制剂在P1位置具有一个与S1特异性位点结合的胍基哌啶基。凝血酶的抑制动力学从慢到快变化,而对于胰蛋白酶,抑制的动力学在所有情况下都快。根据两步机理2中稳定过渡态络合物的缓慢形成来检验动力学。
埃米尔•菲舍尔(Emil Fischer)在1894年提出,酶和底物都具有特定的互补几何形状,彼此恰好契合。这称为“锁和钥匙”模型3。丹尼尔·科什兰(Daniel Koshland)提出了诱导拟合模型,其中底物和酶是相当灵活的结构,当底物与酶4相互作用时,活性位点通过与底物的相互作用不断重塑。
在众多生物活性肽的成熟过程中,需要由其谷氨酰胺(或谷氨酰胺)前体形成N末端焦谷氨酸(pGlu)。游离形式并与底物和三种咪唑衍生抑制剂结合的人QC的结构揭示了类似于两个锌外肽酶的α/β支架,但有多个插入和缺失,特别是在活性位点区域。几种活性位点突变酶的结构分析为针对QC相关疾病5的抑制剂的合理设计提供了结构基础。
作用方式
酶是催化化学反应的蛋白质。酶与底物相互作用并将其转化为产物。抑制剂的结合可以阻止底物进入酶的活性位点和/或阻止酶催化其反应。抑制剂的种类繁多,包括:非特异性,不可逆,可逆-竞争性和非竞争性。可逆抑制剂 以非共价相互作用(例如疏水相互作用,氢键和离子键)与酶结合。非特异性抑制方法包括最终使酶的蛋白质部分变性并因此不可逆的任何物理或化学变化。特定抑制剂 对单一酶发挥作用。大多数毒药通过特异性抑制酶发挥作用。竞争性抑制剂是任何与底物的化学结构和分子几何结构非常相似的化合物。抑制剂可以在活性位点与酶相互作用,但是没有反应发生。非竞争性抑制剂是与酶相互作用但通常不在活性位点相互作用的物质。非竞争性抑制剂的净作用是改变酶的形状,从而改变活性位点,从而使底物不再能与酶相互作用而产生反应。非竞争性抑制剂通常是可逆的。不可逆抑制剂与酶形成牢固的共价键。这些抑制剂可以在活性位点附近或附近起作用。
功能
工业应用中, 酶在商业上被广泛使用,例如在洗涤剂,食品和酿造工业中。蛋白酶用于“生物”洗衣粉中,以加速蛋白质在诸如血液和鸡蛋等污渍中的分解。商业上使用酶的问题包括:它们是水溶性的,这使得它们难以回收,并且一些产物可以抑制酶的活性(反馈抑制)。
药物分子,许多药物分子都是酶抑制剂,药用酶抑制剂通常以其特异性和效力为特征。高度的特异性和效力表明该药物具有较少的副作用和较低的毒性。酶抑制剂在自然界中发现,并且也作为药理学和生物化学的一部分进行设计和生产6。
天然毒物 通常是酶抑制剂,已进化为保护植物或动物免受天敌的侵害。这些天然毒素包括一些已知最剧毒的化合物。
神经气体( 例如二异丙基氟磷酸酯(DFP))通过与丝氨酸的羟基反应生成酯,从而抑制了乙酰胆碱酯酶的活性位点。
参考
1、Scapin G (2006). Structural biology and drug discovery. Curr. Pharm. Des., 12(17):2087–2097.
2、Krishnan R, Zhang E, Hakansson K, Arni RK, Tulinsky A, Lim-Wilby MS, Levy OE, Semple JE, Brunck TK (1998). Highly selective mechanism-based thrombin inhibitors: structures of thrombin and trypsin inhibited with rigid peptidyl aldehydes. Biochemistry, 37 (35):12094-12103.
3、Fischer E (1894). Einfluss der configuration auf die wirkung der enzyme. Ber. Dt. Chem. Ges., 27:2985–2993.
4、Koshland DE (1958). Application of a theory of enzyme specificity to protein synthesis. PNAS., 44 (2):98–104.
5、Huang KF, Liu YL, Cheng WJ, Ko TP, Wang AH (2005). Crystal structures of human glutaminyl cyclase, an enzyme responsible for protein N-terminal pyroglutamate formation. PNAS., 102(37):13117-13122.
6、Holmes CF, Maynes JT, Perreault KR, Dawson JF, James MN (2002). Molecular enzymology underlying regulation of protein phosphatase-1 by natural toxins. Curr Med Chem., 9(22):1981-1989.
Definition
Enzymes are very efficient catalysts for biochemical reactions. They speed up reactions by providing an alternative reaction pathway of lower activation energy. Enzyme acts on substrate and gives rise to a product. Some substances reduce or even stop the catalytic activities of enzymes are called inhibitors.
Discovery
In 1965, Umezawa H analysed enzyme inhibitors produced by microorganisms and isolated leupeptin and antipain inhibiting trypsin and papain, chymostatin inhibiting chymotrypsin, pepstatin inhibiting pepsin, panosialin inhibiting sialidases, oudenone inhibiting tyrosine hydroxylase, dopastin inhibiting dopamine 3-hydroxylase, aquayamycin and chrothiomycin inhibiting tyrosine hydroxylase and dopamine J3-hydroxylase . Recently, an alternative approach has been applied to predict new inhibitors: rational drug design uses the three-dimensional structure of an enzyme's active site to predict which molecules might be inhibitors 1. Computer-based methods for identifying inhibitor for an enzyme have been developed, such as molecular mechanics and molecular docking.
Structural Characteristics
The crystal structures of many inhibitors have been determined. The crystal structures of three highly potent and selective low-molecular weight rigid peptidyl aldehyde inhibitors complexed with thrombin have been determined. All the three inhibitors have a novel lactam moiety at the P3 position, while the two with greatest trypsin selectivity have a guanidinopiperidyl group at the P1 position that binds in the S1 specificity site. The kinetics of inhibition vary from slow to fast with thrombin and are fast in all cases with trypsin. The kinetics are examined in terms of the slow formation of a stable transition-state complex in a two-step mechanism 2.
Emil Fischer in 1894 suggested that both the enzyme and the substrate possess specific complementary geometric shapes that fit exactly into one another.This is known as "the lock and key" model 3. Daniel Koshland suggested induced fit model where substrate and enzymes are rather flexible structures, the active site is continually reshaped by interactions with the substrate as the substrate interacts with the enzyme 4.
N-terminal pyroglutamate (pGlu) formation from its glutaminyl (or glutamyl) precursor is required in the maturation of numerous bioactive peptides. The structure of human QC in free form and bound to a substrate and three imidazole-derived inhibitors reveals an alpha/beta scaffold akin to that of two-zinc exopeptidases but with several insertions and deletions, particularly in the active-site region. The structural analyses of several active-site-mutant enzymes provide a structural basis for the rational design of inhibitors against QC-associated disorders 5.
Mode of Action
Enzymes are proteins that catalyze chemical reactions. Enzymes interact with substrate and convert them into products. Inhibitor binding can stop a substrate from entering the enzyme's active site and/or hinder the enzyme from catalyzing its reaction. There are a variety of types of inhibitors including: nonspecific, irreversible, reversible - competitive and noncompetitive. Reversible inhibitors bind to enzymes with non-covalent interactions like hydrophobic interactions, hydrogen bonds, and ionic bonds. Non-specific methods of inhibition include any physical or chemical changes which ultimately denature the protein portion of the enzyme and are therefore irreversible. Specific Inhibitors exert their effects upon a single enzyme. Most poisons work by specific inhibition of enzymes. A competitive inhibitor is any compound which closely resembles the chemical structure and molecular geometry of the substrate. The inhibitor may interact with the enzyme at the active site, but no reaction takes place. A noncompetitive inhibitor is a substance that interacts with the enzyme, but usually not at the active site. The net effect of a non competitive inhibitor is to change the shape of the enzyme and thus the active site, so that the substrate can no longer interact with the enzyme to give a reaction. Non competitive inhibitors are usually reversible. Irreversible Inhibitors form strong covalent bonds with an enzyme. These inhibitors may act at, near, or remote from the active site .
Functions
Industrial application, enzymes are widely used commercially, for example in the detergent, food and brewing industries. Protease enzymes are used in 'biological' washing powders to speed up the breakdown of proteins in stains like blood and egg. Problems using enzymes commercially include: they are water soluble which makes them hard to recover and some products can inhibit the enzyme activity (feedback inhibition) .
Drug molecules, many drug molecules are enzyme inhibitors and a medicinal enzyme inhibitor is usually characterized by its specificity and its potency. A high specificity and potency suggests that a drug will have fewer side effects and less toxic. Enzyme inhibitors are found in nature and are also designed and produced as part of pharmacology and biochemistry 6.
Natural poisons are often enzyme inhibitors that have evolved to defend a plant or animal against predators. These natural toxins include some of the most poisonous compounds known.
Nerve gases such as diisopropylfluorophosphate (DFP) inhibit the active site of acetylcholine esterase by reacting with the hydroxyl group of serine to make an ester.
References
Scapin G (2006). Structural biology and drug discovery. Curr. Pharm. Des., 12(17):2087–2097.
Krishnan R, Zhang E, Hakansson K, Arni RK, Tulinsky A, Lim-Wilby MS, Levy OE, Semple JE, Brunck TK (1998). Highly selective mechanism-based thrombin inhibitors: structures of thrombin and trypsin inhibited with rigid peptidyl aldehydes. Biochemistry, 37 (35):12094-12103.
Fischer E (1894). Einfluss der configuration auf die wirkung der enzyme. Ber. Dt. Chem. Ges., 27:2985–2993.
Koshland DE (1958). Application of a theory of enzyme specificity to protein synthesis. PNAS., 44 (2):98–104.
Huang KF, Liu YL, Cheng WJ, Ko TP, Wang AH (2005). Crystal structures of human glutaminyl cyclase, an enzyme responsible for protein N-terminal pyroglutamate formation. PNAS., 102(37):13117-13122.
Holmes CF, Maynes JT, Perreault KR, Dawson JF, James MN (2002). Molecular enzymology underlying regulation of protein phosphatase-1 by natural toxins. Curr Med Chem., 9(22):1981-1989.
淀粉肽背景:β淀粉样蛋白(Aβ或Abeta)是从淀粉样前体蛋白加工而成的含有36–43个氨基酸的多肽。Aβ是与阿尔兹海默病相关的淀粉样蛋白斑的成分。已有证据表明,Aβ是一个多功能肽,具有显著的非病理性活性。Aβ是阿尔兹海默病患者脑中发现的沉积物的主要成分。在散发性阿尔兹海默病患者的脑中,Aβ的水平升高,造成脑血管病变和神经毒性。Aβ蛋白是由β和γ分泌酶的连续作用而产生的。γ分泌酶产生Aβ肽的C末端,在APP的转膜结构域切割,可以产生许多36-43个氨基酸残基长度的异构体,最常见的异构体是Aβ40和Aβ42。更长形式的Aβ在内质网中切割产生,而更短形式的Aβ在反面高尔基网中产生。
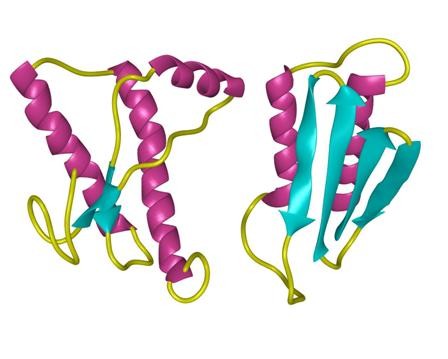
structure of Amyloid β-Peptide (1-40) (human)
淀粉样蛋白肽的 定义淀粉样蛋白 是丝状蛋白质沉积物,大小从纳米到微米不等,并且由肽β链的平行或反平行排列形成的聚集的肽β折叠构成。
结构特征:使用固态NMR(SSNMR),与计算能量最小化过程结合,Tycko和合作者已经提出从淀粉状蛋白肽SS(Aß1-40)的40个残基的形式形成的淀粉样蛋白原纤维的结构在pH 7.4和24 o C在静止条件下。在这种结构中,每个Aß1-40分子在原纤维的核心区域贡献一对ß链,大约跨越残基12-24和30-40。这些由回路25-29连接的链不是同一张ß-sheet的一部分,但参与同一原丝内两个不同的ß-sheets的形成。不同的Aß分子2、3至少从第9到39位残基以平行排列和对齐的方式相互堆叠。通过调用其他实验约束,例如使用透射电子显微镜(TEM)观察到的原丝直径和单位质量通过扫描透射电子显微镜(STEM)1、2测得的长度表明,单个原丝是由四个ß片组成的,它们之间的距离约为10Å。
作用模式:阿尔茨海默氏病(AD)是淀粉样蛋白丝状沉积物的结果,淀粉状蛋白沉积物在分子水平上定义该疾病,发生在神经周膜,轴突,树突和神经元末端,如神经原纤维缠结(NFT),在细胞外神经纤维中淀粉样斑块(APC),以及周围的血管称为淀粉样嗜血性血管病(ACA)。淀粉样蛋白沉积物显然发生在发展NFT的神经元末端区域。已经表明,APC和ACA的主要成分已被证明是4.5kDa的淀粉样蛋白,最初被称为“β-蛋白”或“淀粉样蛋白A4”,我们现在将其称为“βA4”。
功能:钙失调和膜破坏是可溶性淀粉样蛋白低聚物普遍存在的神经毒性机制:进行了一项研究,以研究Ca 2+信号转导可能参与淀粉样蛋白诱导的细胞毒性,疾病相关淀粉样蛋白(β,病毒,胰岛淀粉样蛋白)的均质制剂制备了处于各种聚集状态的多肽,聚谷氨酰胺和溶菌酶),并测试了它们对加载fluo-3的SH-SY5Y细胞的作用。寡聚形式的所有淀粉样蛋白的应用(0.6-6 µg / ml)迅速(约5 s)使细胞内Ca 2+升高,而等量的单体和原纤维则没有。细胞内Ca 2+耗尽后,Abeta42低聚物引起的Ca 2+信号持续存在店,和小信号仍留在钙2 + -游离介质,指示从细胞外和细胞内Ca贡献2+源。膜对Ca 2+的渗透性增加不能归因于内源性Ca 2+通道的活化,因为反应不受强力的Ca 2 +-通道阻滞剂钴的影响。取而代之的是,观察到Abeta42和其他低聚物引起阴离子荧光染料的快速细胞泄漏,这表明膜通透性普遍提高。导致的离子和分子通量失调可能为许多淀粉样变性疾病中Ca 2+失调提供了由低聚物介导的毒性的常见机制。离子起着至关重要的作用,因为它们的跨膜浓度梯度很强,并且参与了细胞功能障碍和死亡。
2型糖尿病中的胰岛淀粉样蛋白和毒性低聚物假说: 2型糖尿病(T2DM)的特征是胰岛素抵抗,胰岛素分泌缺陷,β细胞量减少,β细胞凋亡增加和胰岛淀粉样蛋白。胰岛淀粉样蛋白源自胰岛淀粉样蛋白多肽(IAPP,胰岛淀粉样多肽),该蛋白是通过胰β细胞与胰岛素共表达和共分泌的蛋白。与其他淀粉样蛋白一样,IAPP具有形成膜渗透性毒性低聚物的倾向。越来越多的证据表明,这些有毒的寡聚体而不是这些蛋白质的细胞外淀粉样蛋白形式,是导致神经退行性疾病中神经元丢失的原因。有人提出,胞内IAPP寡聚物的形成可能会导致T2DM 6中的β细胞丢失。
| DOI | 名称 | |
|---|---|---|
| 10.1038/nm754 | Activation of Notch-1 signaling maintains the neoplastic phenotype in human Ras-transformed cells | 下载 |




















