| 编号: | 430043 |
| 中文名称: | Anthranilyl-HIV Protease Substrate VI |
| 英文名: | Anthranilyl-HIV Protease Substrate VI |
| 英文同义词: | ABZ-ARG-VAL-NLE-P-NITRO-PHE-GLU-ALA-NLE-NH2 |
| CAS号: | 210644-49-8 |
| 单字母: | Abz-RV-Nle-F(4NO2)-EA-Nle-NH2 |
| 三字母: | Abz N端Abz修饰 -Arg精氨酸 -Val缬氨酸 -Nle正亮氨酸 -Phe(4NO2)对硝基苯丙氨酸 -Glu谷氨酸 -Ala丙氨酸 -Nle正亮氨酸 -NH2C端酰胺化 |
| 氨基酸个数: | 7 |
| 分子式: | C47H71N13O12 |
| 平均分子量: | 1010.15 |
| 精确分子量: | 1009.53 |
| 等电点(PI): | 8.3 |
| pH=7.0时的净电荷数: | - |
| 平均亲水性: | -0.3 |
| 疏水性值: | 1.2 |
| 消光系数: | - |
| 标签: | 氨基酸衍生物肽 酶底物肽(Substrate Peptide) 侧链保护基肽 人类免疫缺陷病(HIV)肽 |
Caspase酶对应的底物,Caspases(半胱氨酸天冬氨酸蛋白酶,半胱氨酸依赖性天冬氨酸定向蛋白酶)是一类蛋白酶家族,其功能与凋亡(程序性细胞死亡),坏死和发烧(炎症)的过程密切相关。
什么是胱天蛋白酶?
胱天蛋白酶(Caspases)是含半胱氨酸的天冬氨酸蛋白水解酶,它们是为细胞凋亡的主要介质。多种受体,例如TNF-α 受体,FasL受体,TLR和死亡受体,以及Bcl-2和凋亡抑制剂(IAP)蛋白家族参与并调节该caspase依赖性凋亡途径。一旦Caspase受到上游信号(外部或内在)刺激被激活,即会参与执行下游蛋白底物的水解作用,并触发一系列事件,导致细胞分解,死亡,吞噬作用和细胞碎片的清除。
人Caspases酶
人的Caspases家族基于序列相似性和生物学功能等共性主要可分为三大类:第一类由具有长胱天蛋白酶募集结构域的“炎症”胱天蛋白酶组成,他们对P4位上的较大的芳香族或疏水性残基具有亲和力。第二类由具有短的前体结构域的“细胞凋亡效应”胱天蛋白酶组成,而第三类由具有长的前提结构域的Pap位置具有亮氨酸或缬氨酸底物亲和力的“凋亡引发剂”胱天蛋白酶组成(表1)。
表1. 人胱天蛋白酶的功能分类:
| 细胞死亡途径 | 半胱天冬酶类型 | 酵素 | 物种 |
| 细胞凋亡 | 启动器 | Caspases 2 | 人与鼠 |
| 细胞凋亡 | 启动器 | Caspases 8 | 人与鼠 |
| 细胞凋亡 | 启动器 | Caspases 9 | 人与鼠 |
| 细胞凋亡 | 启动器 | Caspases 10 | 人的 |
| 细胞凋亡 | 效应器 | Caspases 3 | 人与鼠 |
| 细胞凋亡 | 效应器 | Caspases 6 | 人与鼠 |
| 细胞凋亡 | 效应器 | Caspases 6 | 人与鼠 |
| 细胞焦亡 | 炎性的 | Caspases 1 | 人与鼠 |
| 细胞焦亡 | 炎性的 | Caspases 4 | 人的 |
| 细胞焦亡 | 炎性的 | Caspases 5 | 人的 |
启动器Caspase和效应器Caspase酶
根据其在凋亡胱天蛋白酶途径中的作用,胱天蛋白酶可分为两类:启动器和效应器Caspase酶。启动器和效应器Caspas酶都具有由小亚基和大亚基组成的催化位点,Caspase酶的识别位
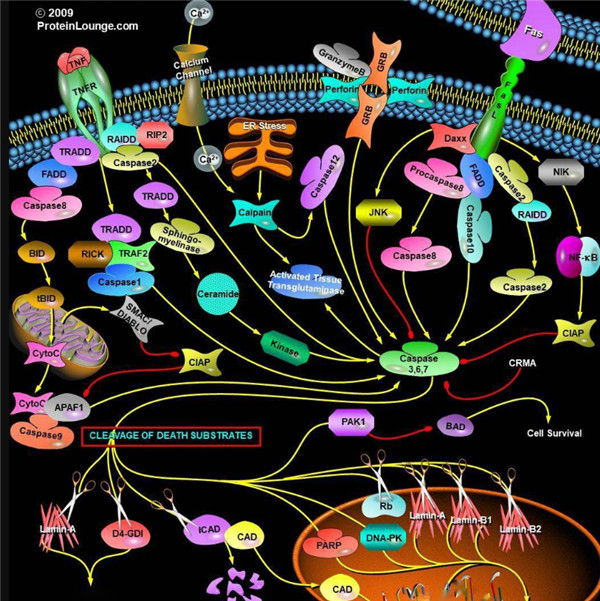
凋亡启动器Caspase酶,例如caspase-2,-8,-9和-10可以启动caspase激活级联反应。Caspase-8对于形成死亡诱导信号复合物(DISC)是必不可少的,并且在激活后,Caspase-8激活下游效应子Caspase(例如Caspase 3)并介导线粒体中细胞色素c的释放。Caspase-8已被证明对IETD肽序列具有相对较高的底物选择性。凋亡效应胱天蛋白酶例如Caspase-3,-6和-7虽然不负责启动级联途径,但是当被激活时,它们在级联的中间和后续步骤中起着不可或缺的作用。Caspase-3(CPP32 / apopain)是关键效应器,因为它放大了来自启动器Caspase的信号,使用对Caspase-3有选择性的DEVD肽序列对活化的Caspase-3进行检测,可以检测Caspase-3的活性。
Caspase酶底物和抑制剂
Caspase底物和抑制剂由两个关键成分组成:Caspase识别序列和信号产生或蛋白酶抑制基序。不同Caspase识别序列不同,一般由三个或四个氨基酸组成(表2)。Caspase酶识别序列的N端通常有乙酰基(Ac)或碳苯甲氧基(Z)基团修饰,以增强膜的通透性。对应的Caspase识别特定的肽序列为其酶促反应切割位点,释放产生信号或抑制信号的基序。Caspase的显色和荧光底物均以相似的方式起作用,其中底物的信号或颜色强度与蛋白水解活性成正比。
表2. Caspase的底物及其序列
| 多肽 | 氨基酸序列 | 对应的Caspase的种类 |
| IETD | Ile-Glu-Thr-Asp | Caspase 8,颗粒酶B |
| DEVD | Asp-Glu-Val-Asp | Caspase 3、6、7、8或10 |
| LEHD | Leu-Glu-His-Asp | Caspase 9 |
| VAD | Val-Ala-Asp | Caspase 1、2、3、6、8、9或10 |
Caspase酶的显色底物
Caspase的显色底物是有Caspase识别序列及生色基团组成,常见的生色团有pNA(对硝基苯胺或4-硝基苯胺),可使用酶标仪或分光光度计在405 nm处进行光密度检测。
表3. Caspase的显色底物
| 底物 | Caspase | 吸收(nm) | 颜色 |
| Ac-DEVD-pNA * CAS 189950-66-1 * | 半胱天冬酶3 | 405 nm | 黄色 |
| Z-DEVD-pNA | 半胱天冬酶3 | 405 nm | 黄色 |
| Z-IETD-pNA * CAS 219138-21-3 * | 半胱天冬酶8,颗粒酶B | 405 nm | 黄色 |
Caspase的荧光底物
Caspase的荧光底物的结构包含与半胱天冬酶识别相关的荧光团,例如7-氨基-4-甲基香豆素(AMC),7-氨基-4-三氟甲基香豆素(AFC), Rhodamine 110(R110)或ProRed™620。R110的Caspase底物比基于香豆素的Caspase底物(例如AMC和AFC)更敏感,但由于两步裂解过程,其动态范围更窄。 建议将R110标记的Caspase底物用于终点法测定,而将AMC和AFC标记的 Caspase底物用于动力学测定。
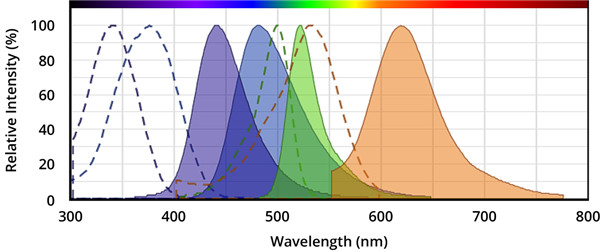
图.从左到右,分别是AMC(7-氨基-4-甲基香豆素),AFC(7-氨基-4-三氟甲基香豆素),Rhodamine 110(R110)和ProRed™620的激发和发射光谱。
表4.荧光半胱天冬酶底物。
| 底物名称 | 对应的Caspase | Ex(nm) | Em(nm) | ε¹ | Φ² |
| Ac-DEVD-AFC * CAS 201608-14-2 * | 半胱天冬酶3、7 | 376 | 482 | 17000 | 0.53 |
| Ac-DEVD-AMC * CAS 169332-61-0 * | 半胱天冬酶3、7 | 341 | 441 | 19000 | N / D |
| Z-DEVD-AFC | 半胱天冬酶3、7 | 376 | 482 | 17000 | 0.53 |
| Z-DEVD-AMC * CAS 1135416-11-3 * | 半胱天冬酶3、7 | 341 | 441 | 19000 | N / D |
| Z-DEVD-ProRed™620 | 半胱天冬酶3、7 | 532 | 619 | N / D | N / D |
| (Z-DEVD)2 -R110 * CAS 223538-61-2 * | 半胱天冬酶3、7 | 500 | 522 | 80000 | N / D |
| Z-DEVD-ProRed™620 | 半胱天冬酶3、7 | 532 | 619 | N / D | N / D |
| Ac-IETD-AFC * CAS 211990-57-7 * | 半胱天冬酶8,颗粒酶B | 376 | 482 | 17000 | 0.53 |
| Z-IETD-AFC * CAS 219138-02-0 * | 半胱天冬酶8,颗粒酶B | 376 | 482 | 17000 | 0.53 |
注意:
1.ε=在其最大吸收波长处的摩尔消光系数(单位= cm -1M -1)。
2.Φ=水性缓冲液(pH 7.2)中的荧光量子产率。
Caspase抑制剂
Caspase抑制剂能与Caspase的活性位点结合并形成可逆或不可逆的连接,通常,Caspase抑制剂的结构由Caspase识别序列,诸如醛(-CHO)或氟甲基酮(-FMK)的官能团组成。具有醛官能团的胱天蛋白酶抑制剂是可逆的,而具有FMK的抑制剂是不可逆的。半胱天冬酶底物和抑制剂都具有较小的细胞毒性作用,因此,它们是研究半胱天冬酶活性的有用工具。
表5. 可逆和不可逆的Caspase酶抑制剂
| 抑制剂 | Caspase的种类 | 是否可逆 | Ex(nm) | Em(nm) |
| Ac-DEVD-CHO * CAS 169332-60-9 * | 半胱天冬酶3、7 | 可逆的 | -- | -- |
| Ac-IETD-CHO * CAS 191338-86-0 * | 半胱天冬酶8 | 可逆的 | -- | -- |
| mFluor™450-VAD-FMK | 半胱天冬酶1,2,3,6,8,9,10 | 不可逆的 | 406 | 445 |
| mFluor™510-VAD-FMK | 半胱天冬酶1,2,3,6,8,9,10 | 不可逆的 | 412 | 505 |
| FITC-C6-DEVD-FMK | 半胱天冬酶3、7 | 不可逆的 | 491 | 516 |
| FITC-C6-DEVD-FMK | 半胱天冬酶3、7 | 不可逆的 | 491 | 516 |
| FITC-C6-LEHD-FMK | 半胱天冬酶9 | 不可逆的 | 491 | 516 |
| FITC-C6-LEHD-FMK | 半胱天冬酶9 | 不可逆的 | 491 | 516 |
| FAM-VAD-FMK | 半胱天冬酶1,2,3,6,8,9,10 | 不可逆的 | 493 | 517 |
| SRB-VAD-FMK [磺胺丁胺B-VAD-FMK] | 半胱天冬酶1,2,3,6,8,9,10 | 不可逆的 | 559 | 577 |
Definition
Human immunodeficiency virus (HIV) is a lentivirus that causes acquired immunodeficiency syndrome (AIDS), a condition in humans in which the immune system begins to fail, leading to life-threatening opportunistic infections. Over 5000 HIV-related peptides have been synthesized, that inhibit different stages of viral life cycle.
Discovery
In 1983, two separate researchers Robert Gallo and Luc Montagnier independently declared that a novel retrovirus infecting AIDS patients. Several HIV related peptides including peptides (15-mers or 20-mers) of HIV glycoprotein 160 (gp160), gp120W16D, MN envelope (env) consensus B tat, consensus B VIF, HXB2 gag, SIVmac239, SIVmac239env, SIVmac239 gag have been used to study HIV life cycle. C34 peptide of Gp41 HIV Fragment is known as HR2, belongs to the helical region of gp41 of HIV, C-terminal heptad repeat 2 (HR2) defined as C helix or C peptide. It is known that HIV-1 enters cells by membrane fusion, C34 gp41 peptide is a potent inhibitors of HIV-1 fusion 1,2. The 86 amino acid trans-activator (Tat) protein of human immunodeficiency virus type 1 (HIV-1) is an RNA-binding transcriptional regulator. HIV-1 Tat proteins (wild type and Thr40Lys mutant) and the HIV-1 Tat peptide fragments Tat(32–48) and Tat(32–72) were chemically synthesized and used for HIV studies 3.
HIV (gp120) fragment (254-274), this fragment with sequence homology to a domain of the external envelope glycoprotein (gp120) of the human immunodeficiency virus (HIV) is important for HIV infectivity and antibody neutralization 4. HIV (gp120) fragment (421-438), derived from the CD4 attachment region of HIV gp120, inhibited the syncytial formation in vitro 5. HIV-1 gag protein p17 (76-84), HLA-A*0201-restricted immunodominant CD8 epitope of the HIV gag protein used for the characterization of CD8+ -T cells of HIV-positiv patients 6. HIV-1 rev protein (34-50), this arginine-rich fragment interacts specifically with RNA. It has been shown that rev protein and rev protein (34-50) bind IIB RNA with a similar dissociation constant of approx. 10 nM 7.
Structural Characteristics
The HIV type-1 belongs to the family Retroviridae and consists of two basic components: a core of ribonucleic acid (RNA), called the genome, and a protein component that surrounds the genome, called a capsid. The single-stranded RNA is tightly bound to nucleocapsid proteins and enzymes needed for the morphogenesis of the virion such as reverse transcriptase, proteases, ribonuclease and integrase. A matrix composed of the viral protein that surrounds the capsid. Viral envelope is composed of two layers of fatty molecules taken from the membrane of a human cell during budding process. There are 70 copies of a complex HIV protein that protrudes through the surface of the virus particle, known as Env, consists of a cap, glycoprotein (gp) 120, and a stem, gp41 molecules. This glycoprotein complex is important for fusion of virus to host cell. Both these surface proteins are important targets for treatments or HIV vaccines 8.
Mode of Action
HIV binds to a CD4 receptor and one of two co-receptors on the surface of a CD4+ T- lymphocyte. After fusion, the virus releases RNA, its genetic material, into the host cell. An HIV enzyme called reverse transcriptase converts the single- stranded HIV RNA to double-stranded HIV DNA. The newly formed HIV DNA enters the host cell's nucleus. The integrated HIV DNA is called provirus. The provirus may remain inactive for several years, producing few or no new copies of HIV. When the host cell receives a signal to become active, the provirus uses a host enzyme called RNA polymerase to create copies of the HIV genomic material, as well as shorter strands of RNA called messenger RNA (mRNA). The mRNA is used as a blueprint to make long chains of HIV proteins. An HIV enzyme called protease cuts the long chains of HIV proteins into smaller individual proteins. As the smaller HIV proteins come together with copies of HIV's RNA genetic material, a new virus particle is assembled. The newly assembled virus buds out from the host cell. During budding, the new virus acquires part of the cell's outer envelope. This envelope is embedded with viral glycoproteins which are necessary for host cell recognition.
Functions
CD8 cytotoxic, HIV-1 specific CD8 cytotoxic T lymphocyte (CTL) responses play a critical role in controlling HIV-1 replication. TCR avidity correlates with CTL function, and CTLs expressing TCRs with high avidity for their cognate MHC-viral peptide complex play an important in vivo role in neutralizing virus infections, terminating virus infection and delaying systemic AIDS virus dissemination from the mucosal inoculation site.
HIV-1 envelope transmembrane protein that contain highly positively charged amphipathic helices (designated LLP) in have both cytolytic and calmodulin (CaM) binding/inhibitory properties that contribute to cytopathogenesis during a viral infection.
HIV-1 vif, The human immunodeficiency virus type 1 (HIV-1) auxiliary gene vif is essential for virus propagation in peripheral blood lymphocytes, macrophages, and in some T-cell lines. (i) Vif protein binds HIV-1 PR (protease), but not covalently linked tethered PR-PR; (ii) the four amino acids residing at the N terminus of HIV-1 PR are essential for Vif/PR interaction; (iii) synthetic peptide derived from the N terminus of HIV-1 PR inhibits Vif/PR binding; and (iv) this peptide inhibits the propagation of HIV-1 in restrictive cells 9.
References
1. Bianchi E, Finotto M, Ingallinella P, Hrin R, Carella AV, Hou XS, Schleif WA, Miller MD, Geleziunas R, Pessi A (2005). Covalent stabilization of coiled coils of the HIV gp41 N region yields extremely potent and broad inhibitors of viral infection. PNAS., 102(36):12903-12908
2. de Rosny E, Vassell R, Jiang S, Kunert R, Weiss CD (2004). Binding of the 2F5 monoclonal antibody to native and fusion-intermediate forms of human immunodeficiency virus type 1 gp41: implications for fusion-inducing conformational changes. J. Virol., 78(5):2627-2631.
3. Klostermeier D, Bayer P, Kraft M, Frank RW, Rösch P (1997). Spectroscopic investigations of HIV-1 trans-activator and related peptides in aqueous solutions. Biophysical Chemistry, 63(2):87-96.
4. Ho DD, Kaplan JC, Rackauskas IE, Gurney ME (1988). Second conserved domain of gp120 is important for HIV infectivity and antibody neutralization. Science, 239(4843):1021-1023.
5. Morrow WJ, Williams WM, Whalley AS, Ryskamp T, Newman R, Kang CY, Chamat S, Köhler H, Kieber-Emmons T (1992). Synthetic peptides from a conserved region of gp120 induce broadly reactive anti-HIV responses. Immunology, 75(4):557-564.
6. Wilkinson J, Cope A, Gill J, Bourboulia D, Hayes P, Imami N, Kubo T, Marcelin A, Calvez V, Weiss R, Gazzard B, Boshoff C, Gotch F (2002). Identification of Kaposi's sarcoma-associated herpesvirus (KSHV)-specific cytotoxic T-lymphocyte epitopes and evaluation of reconstitution of KSHV-specific responses in human immunodeficiency virus type 1-Infected patients receiving highly active antiretroviral therapy. J. Virol., 76(6):2634-2640.
7. Kjems J, Calnan BJ, Frankel AD, Sharp PA (1992). Specific binding of a basic peptide from HIV-1 Rev. EMBO J., 11(3):1119-29.
8. Chan DC, Fass D, Berger JM, Kim PS (1997). Core structure of gp41 from the HIV envlope glycoprotein . Cell, 89:263–73.
9. Hutoran M, Britan E, Baraz L, Blumenzweig I, Steinitz M, Kotler M (2004). Abrogation of Vif function by peptide derived from the N-terminal region of the human immunodeficiency virus type 1 (HIV-1) protease. Virology, 330(1):261-270.




















